Salzig (Typ I - Zellen)
Alle Tiere haben Geschmacksrezeptoren für die Geschmacksqualität "salzig". Die Aufnahme von Salzen ist wichtig für die Aufrechterhaltung des Elektrolythaushalts [1].
Geschmacksrezeptoren, die auf die Geschmacksqualität "salzig" reagieren, sind relativ einfach aufgebaut. Kochsalz enthält bekanntlich Natrium-Ionen, und wenn Na+-Ionen in eine Zelle eindringen, erhöht sich deren Membranpotenzial teils erheblich (Depolarisation). Eine solche Depolarisation wandert dann wellenförmig (zeitliche und räumliche Abschwächung) zum basalen Ende der Sinneszelle, wo dann gegebenenfalls Neurotransmitter ausgeschüttet werden, die die nachfolgende Nervenzelle erregen.
Nach diesem "Schulbuchwissen" wollen wir uns nun mal die Einzelheiten dieses Transduktionsvorgangs näher anschauen.
Einzelheiten
Die Natriumkanäle der Salzig-Rezeptoren sind weder chemisch noch elektrisch gesteuert, sondern ständig offen. Na+-Ionen können leicht durch diese Kanäle in die Zelle eindringen und dort ein Rezeptorpotenzial auslösen. Ist dieses Rezeptorpotenzial am basalen Ende der Zelle hoch genug, öffnen sich dort spannungsgesteuerte Natrium- und Calcium-Kanäle. Die Depolarisation wird dort verstärkt, und die Calcium-Ionen sorgen für die Verschmelzung der präsynaptischen Vesikeln mit der präsynaptischen Membran, so dass die Neurotransmitter ausgeschüttet werden können.
Andere Kationen wie K+, Mg2+ oder Ca2+ können nicht durch die stets offenen Na+-Kanäle im vorderen Ende des Geschmacksrezeptors eindringen, nur Na+-Ionen sorgen für den typisch salzigen Geschmack.
Ist die Na+-Konzentration auf der Zunge zu hoch, wird der salzige Geschmack von sauren und bitteren Empfindungen überlagert, weil bei hohen Na+-Konzentrationen auch die Rezeptoren für diese Geschmacksqualitäten erregt werden. Eine versalzene Suppe schmeckt daher nicht mehr angenehm salzig, sondern unangenehm sauer bis bitter.
Na+-Ionen treten ja nie alleine auf, sondern werden immer von einem passenden Gegen-Anion begleitet. Im Kochsalz ist dies das Chlorid-Ion Cl-. Der Geschmackseindruck "salzig" hängt auch von dem Gegen-Anion ab. Natriumacetat (Gegen-Anion = CH3COO-) schmeckt beispielsweise nicht mehr so salzig wie Natriumchlorid. Offensichtlich hemmt das größere Anion die Rezeptoren. Ob diese Hemmung auf einem einfachen Verdünnungsprozess beruht (die großen Anionen lassen weniger Platz für die kleinen Kationen) oder ob ein komplexerer Mechanismus dahinter steckt, ist noch nicht geklärt [2, S 284].
Sehr große Anionen haben sogar einen eigenen Geschmack, der den Salzig-Geschmack der Natrium-Ionen überdecken kann. Natriumcitrat beispielsweise schmeckt nicht mehr salzig, sondern nach Seife [3].
Sauer (Typ III - Zellen)
Im Prinzip funktionieren die Geschmackssinneszellen für "sauer" ähnlich wie die "salzig"-Rezeptoren. Bei Erregung strömen Calcium-Ionen durch spezielle Kanäle in den basalen Teil der Zellen ein und sorgen für die Freisetzung von Neurotransmittern wie Serotonin, GABA und Noradrenalin [4, 5]. Trotzdem gehören nur die "sauer"-Rezeptoren zu den Typ III - Zellen, während die "salzig"-Rezeptoren zu den Typ I - Zellen gezählt werden.
Einer der Unterschiede zwischen Typ I - und Typ III - Zellen liegt darin, dass Typ III - Zellen h durch das ATP erregt werden können, das von den Typ II - Zellen ausgeschüttet wird, während Typ I - Zellen ATP-unempfindlich sind. Typ III - Zellen sprechen daher indirekt auch auf die vier Geschmacksrichtungen "süß", "umami", "fettig" und "bitter" an.
Zum eigentlichen Transduktionsprozess:
Im Buch von Bear, Connors und Paradiso [2] wird vorgeschlagen, dass einerseits H+-Ionen durch spezielle Ionenkanäle direkt in die Zelle strömen und dort für eine leichte Depolarisierung sorgen. Andererseits sollen die H+-Ionen bestimmte Kalium-Kanäle hemmen, so dass keine K+-Ionen mehr nach außen strömen können. Durch die Hemmung dieser K+-Kanäle wird die Depolarisierung der Membran verstärkt.
Am basalen Ende des "sauer"-Rezeptors laufen dann die gleichen Vorgänge ab wie beim "salzig"-Rezeptor (siehe vorherigen Abschnitt).
Bitter, Süß, Umami, Fett (Typ II - Zellen)
Der Transduktionsprozess der Typ II - Zellen
Diese drei Transduktionsprozesse funktionieren auf die gleiche Weise, allerdings mit unterschiedlichen Rezeptorproteinen für die vier Geschmacksrichtungen [4].
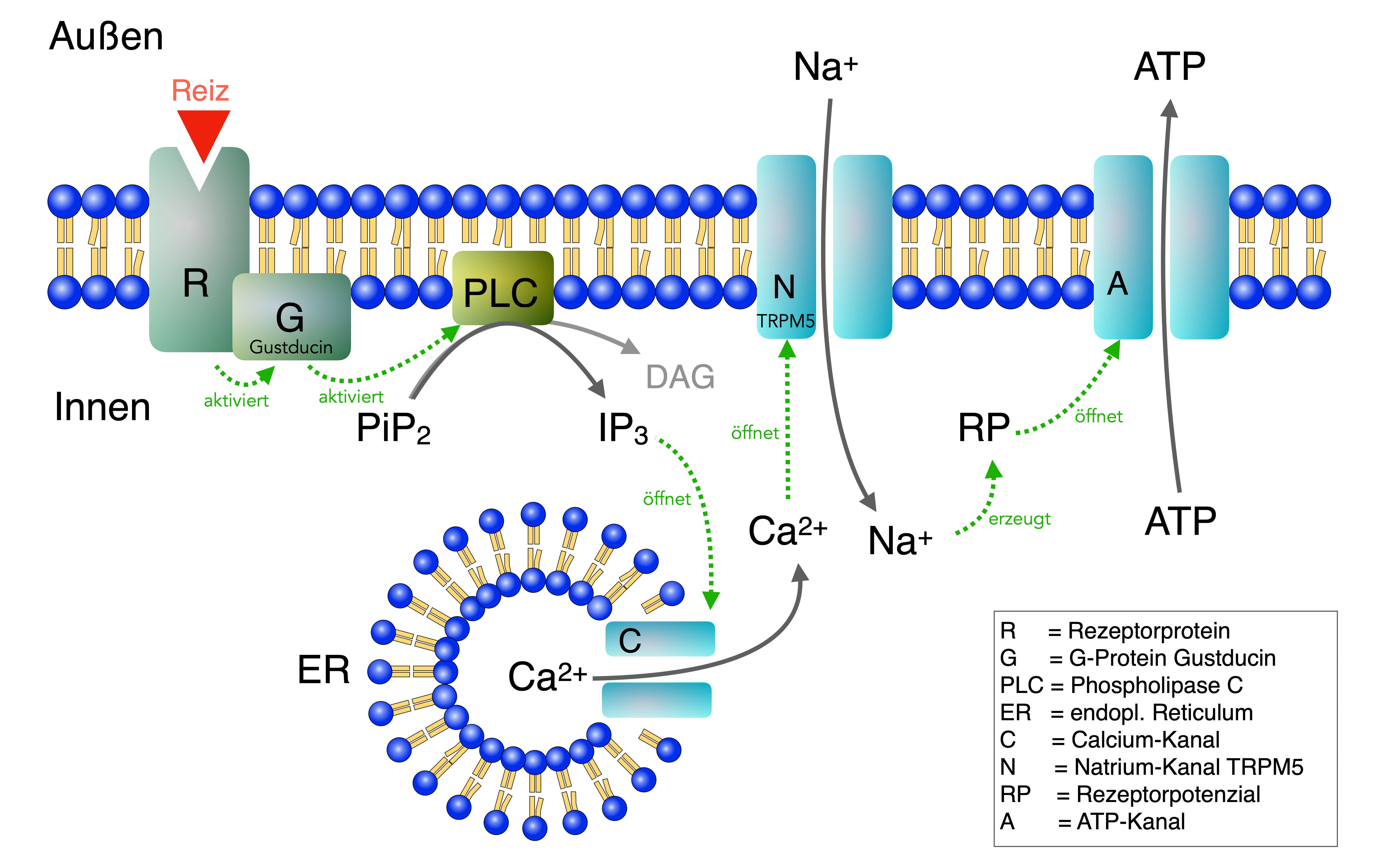
Transduktion bei den Typ II - Rezeptoren, nach [2 und 4]
Autor: Ulrich Helmich 2022, Lizenz: Public domain
Durch Klicken auf das Bild wird eine größere Version geladen
Ein Molekül eines bitteren, süßen, fettigen oder nach Fleisch schmeckenden (umami) Geschmacksstoffs dockt an das Rezeptorprotein (R) in der Cilienmembran der Geschmackssinneszelle an. Dies aktiviert auf der Membraninnenseite ein G-Protein (G) namens Gustducin, welches wiederum eine Phospholipase C (PLC) aktiviert. Dieses Enzym setzt das Membranlipid PiP2 zu den sekundären Botenstoffen IP3 und DAG um.
Das IP3wiederum öffnet Calcium-Kanäle (C) im endoplasmatischen Reticulum (ER), die daraufhin Ca2+-Ionen in das Cytoplasma entlassen. Diese Calcium-Ionen setzen sich an spezifische TRPM5-Natrium-Kanäle (N) und öffnen diese, so dass Na+-Ionen in die Zelle strömen. Dadurch wird die Zellmembran depolarisiert, es entsteht ein Rezeptorpotenzial (RP). Dieses Rezeptorpotenzial führt dann zur Öffnung bestimmter ATP-Kanäle, durch die ATP nach außen strömt und hier wie ein Neurotransmitter wirkt.
Die Skizze in Abbildung 1 ist in dreierlei Hinsicht vereinfacht:
- Erstens muss man sich vorstellen, dass die Rezeptoren (R) mit ihren G-Proteinen (G) und den Phospholipasen (PLC) am oberen Ende der Sinneszellen sitzen, in den Cilien, während die Natriumkanäle (N) und die ATP-Kanäle (A) am basalen unteren Ende der Zellen sitzen.
- Zweitens besteht das Rezeptorprotein (R) aus zwei unterschiedlichen Einheiten, es handelt sich um ein sogenanntes Heterodimer. Der Süß-Rezeptor besteht aus den Proteinen T1R2 und T1R3, der Umami-Rezeptor aus den Proteinen T1R1 und T1R3, der Fett-Rezeptor aus den Einheiten CD36 und GPR120 und der Bitter-Rezeptor schließlich setzt sich aus den Proteineinheiten T2Rx und nochmal T2Rx zusammen, ist also ein Homodimer.
- Drittens aktiviert ein Rezeptor-Dimer mehrere G-Proteine gleichzeitig, und jedes G-Protein kann mehrere Phospholipasen aktivieren. Jede dieser Phospholipasen kann viele IP3-Moleküle herstellen, die entsprechend viele Calcium-Kanäle im ER öffnen. Wenige Geschmacks-Moleküle können also eine ganze Reaktionskaskade in Gang setzen, eine Lawine sozusagen. Daher kann man bestimmte Geschmacksstoffe schon in extrem geringen Konzentrationen schmecken.
Das am Ende freigesetzte ATP wirkt auf die präsynaptischen Membranen der nachfolgenden Nervenzellen wie ein Neurotransmitter, führt also zu einer Depolarisierung derselben. Die Typ II - Geschmackssinneszelle kann ja keine Aktionspotenziale bilden, sie hat noch nicht einmal synaptische Vesikel, sondern schüttet bei Erregung einfach ATP aus.
Weitere Einzelheitem zum Süß-Rezeptor und zur Bedeutung des süßen Geschmacks finden Sie auf dieser Vertiefungsseite.
Weitere Einzelheitem zum Umami-Rezeptor und zur Bedeutung des würzigen Geschmacks finden Sie auf dieser Vertiefungsseite.
25 Rezeptoren für "bitter"
Beim Menschen gibt es ca. 25 verschiedene Rezeptorproteine für die Geschmacksqualität "bitter". Allerdings produziert eine Geschmackssinneszelle für "bitter" sehr viele dieser 25 Rezeptoren, so dass viele verschiedene Bitterstoffe als "bitter" erkannt werden. Eine Unterscheidung zwischen verschiedenen "bitter"-Qualitäten ist allerdings nicht möglich, weil die verschiedenen Rezeptoren in der gleichen Zellmembran sitzen und die gleichen molekularen Vorgänge auslösen. Die hohe Anzahl der "bitter"-Rezeptoren ermöglicht nur, dass möglichst viele chemische Verbindungen erkannt werden, die potenziell giftig sind [2].
Wer sich näher über die "bitter"-Rezeptoren informieren will, kann sich ja mal die Dissertation von Sandra Hübner anschauen [5], sie schreibt über die "Molekularen Grundlagen der Bittergeschmackswahrnehmung in der Maus". Zumindest die Einleitung sollte man lesen!
Auf dieser Seite finden Sie weitere Einzelheiten zu den Bitter-Rezeptoren des Menschen.
Quellen, die über allgemeines Schulbuchwissen hinausgehen:
- Kandel, Schwartz, Jessel, Siegelbaum, Hudspeth, Principles of Neural Science, Fifth Edition. McGraw-Hill Education 2013. Kindle-Version.
- Bear, Connors, Paradiso: Neurowissenschaften, Springer-Verlag 2018
- Wikipedia, Artikel "Natriumcitrat"
- Schling: Der Geschmack (essentials). Springer-Verlag 2018. Kindle-Version.
- Hübner: Molekulare Grundlagen der Bittergeschmackswahrnehmung in der Maus. Dissertation, Uni Potsdam 2014.