Entstehung der Primordialwand
Die Zellwand der Pflanzenzelle wird schon während der Mitose angelegt.
Allgemeine Informationen zur pflanzlichen Zellwand finden Sie hier.
Man kann schon in der Anaphase eine Anhäufung von Golgi-Vesikeln in der Äquatorialebene beobachten:
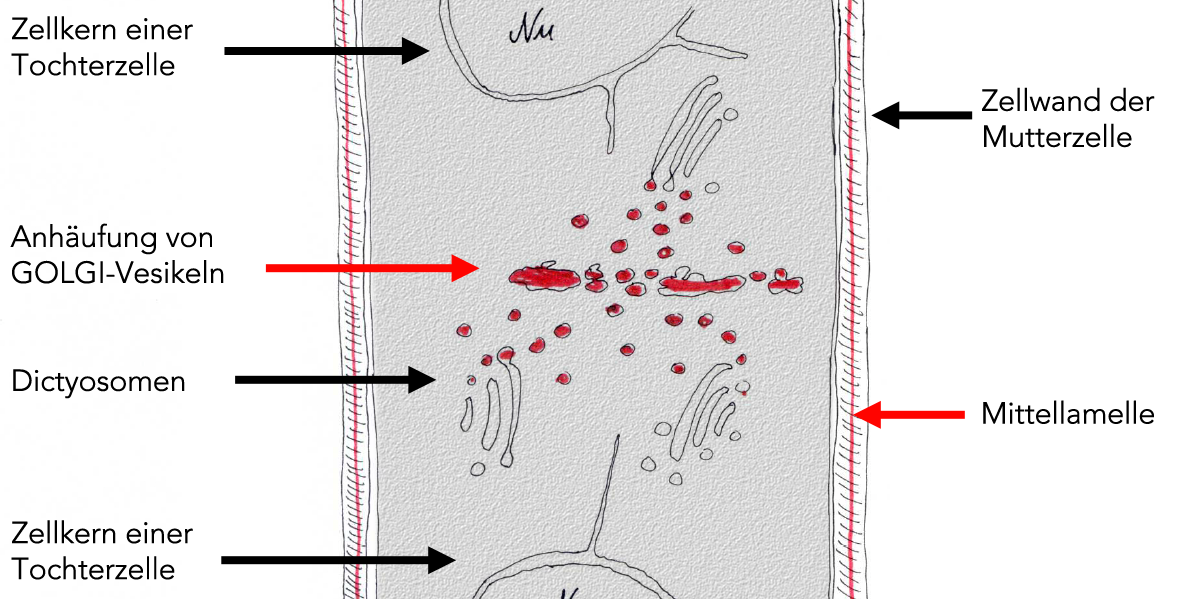
Bildung der Zellplatte aus Golgi-Vesikeln
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Diese Golgi-Vesikel enthalten Polysaccharide, die zum Aufbau der ersten Zellwandschicht dienen. Gleichzeitig enthalten sie Membranmaterial zur Bildung der neuen Plasmamembran der beiden Tochterzellen.
Einzelheiten zum Golgi-Apparat, wie er mit dem ER in Verbindung steht und welche Arten von Golgi-Vesikeln es gibt, erfahren Sie auf dieser Lexikon-Seite.
Der nächste Schritt - in der Abbildung bereits angedeutet - ist die Verschmelzung der einzelnen Golgi-Vesikel. Es entsteht ein Membrangebilde, das Polysaccharide enthält. Als Nächstes vereinigt sich die Membran dieses Gebildes mit der Zellmembran der Mutterzelle.
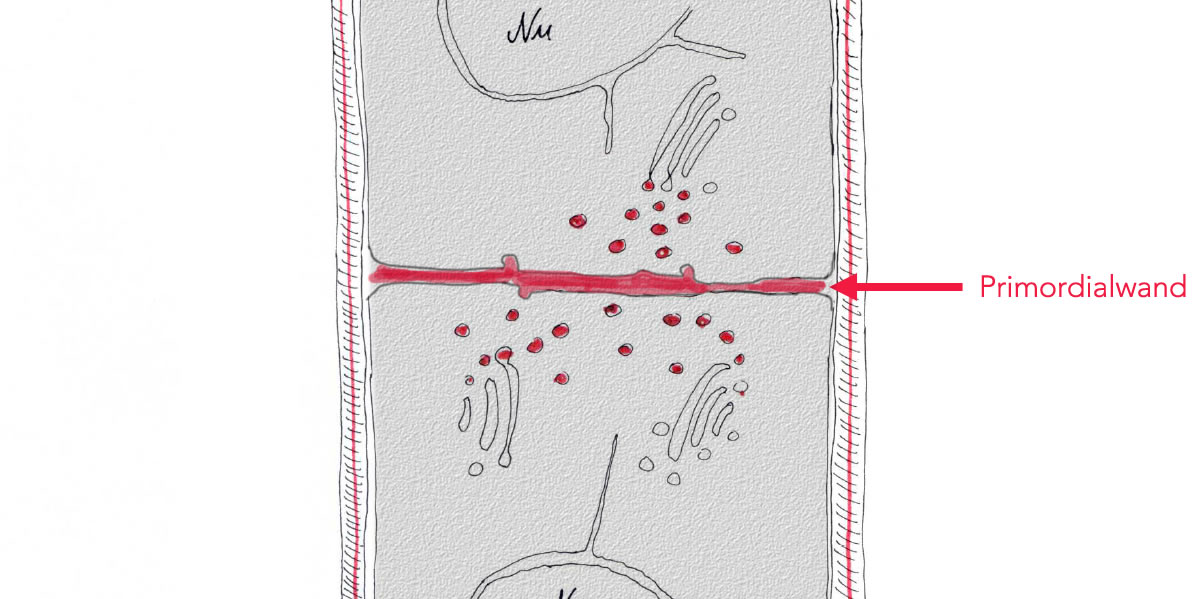
Bildung der Primordialwand
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Dies führt schließlich zur Durchschnürung der Mutterzelle in zwei Tochterzellen.
Gleichzeitig ordnen sich die Polysaccharide so aus, dass eine erste Zellwandschicht, die sogenannte Primordialwand entsteht. Allerdings besteht die Primordialwand nicht als geschlossenes Gebilde, sondern es werden Durchgänge ausgespart, welche die Tochterzellen verbinden:
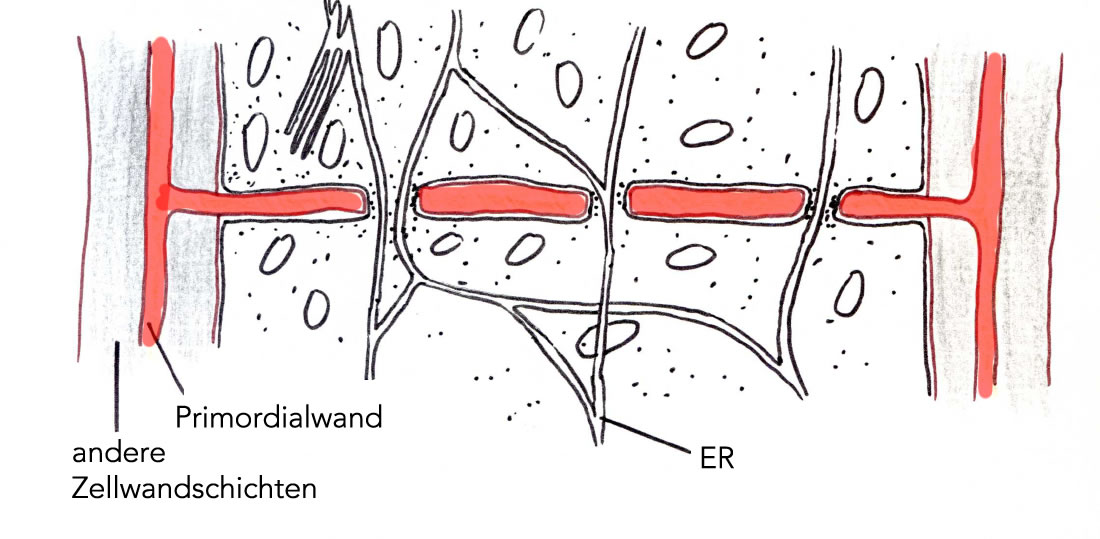
Fortgeschrittenere Bildung der Primordialwand
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Diese Verbindungskanäle werden vom Cytoplasma und der Plasmamembran durchzogen und heißen Plasmodesmen.
Zusammensetzung der Primordialwand
Die Polysaccharide, aus denen die Primordialwand besteht, sind in der Gruppe der Pektine anzuordnen. Im folgenden Bild sehen wir typische Makromoleküle, wie sie im Pektin vorkommen.
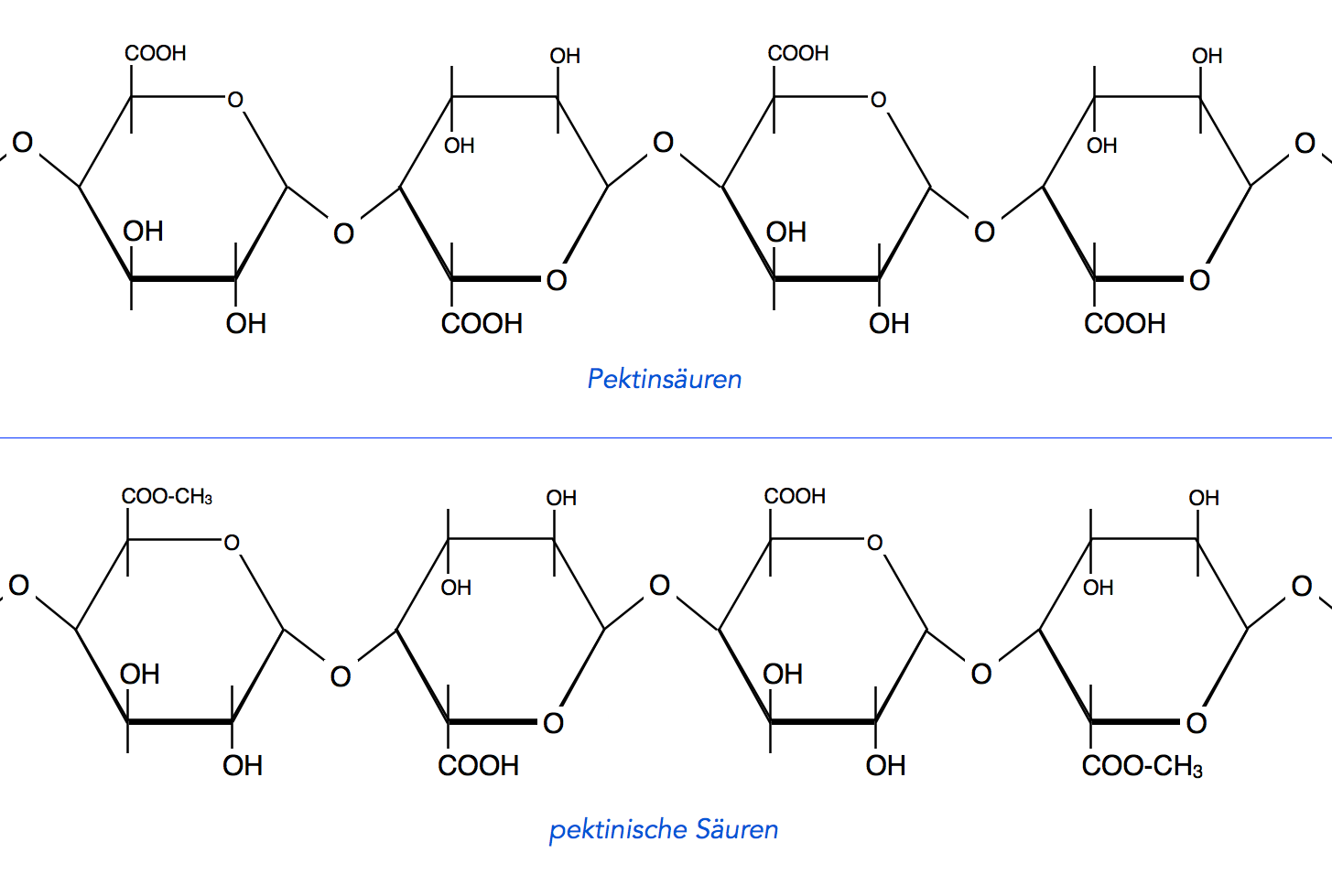
Pektinsäuren und "pektinische Säuren"
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Als Pektinsäuren bezeichnet man Polymere der α-D-Galacturonsäure. Das ist ein Oxidationsprodukt der Galactose, einem Monosaccharid, das auch in Milchzucker enthalten ist.
Der Begriff "pektinische Säuren" ist nicht mehr so oft in Gebrauch; es handelt sich um Pektinsäuren, die zu ca. 50% mit dem Alkohol Methanol CH3-OH verestert sind.
Für Experten:
Die linearen D-Galacturonsäureketten (Pektinsäuren) sind weiterhin durch L-Rhamnose-Einheiten miteinander verbunden:
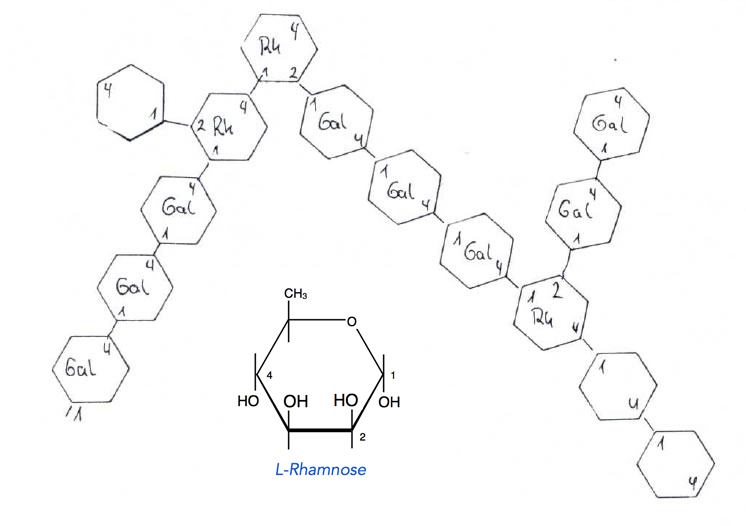
Molekularstruktur eines Pektins
Autor: Ulrich Helmich, Lizenz: Public domain
Die L-Rhamnose-Monomere können D-Galacturonsäureketten über die C-Atome 1, 2 und 4 miteinander verketten. Weiterhin ist die Galacturonylhauptkette mit kurzen Ketten aus neutralen Zuckern wie Galactose, Arabinose, Xylose und Glucose verbunden. Mit Hilfe zweiwertiger Kationen wie Mg2+ oder Ca2+ können die einzelnen Pectinsäure-Moleküle über ihre negativ geladenen COO--Gruppen zu wasserunlöslichen Riesenmolekülen vernetzt werden: Protopectin.
Zu diesem Thema existiert leider noch keine eigene Seite auf dieser Homepage, daher verweise ich hier auf den Artikel in der Wikipedia.
Bildung der Primärwand
Auf diese Primordialwand wird nun durch Golgi-Vesikel eine weitere Schicht aufgelagert, die Primärwand:
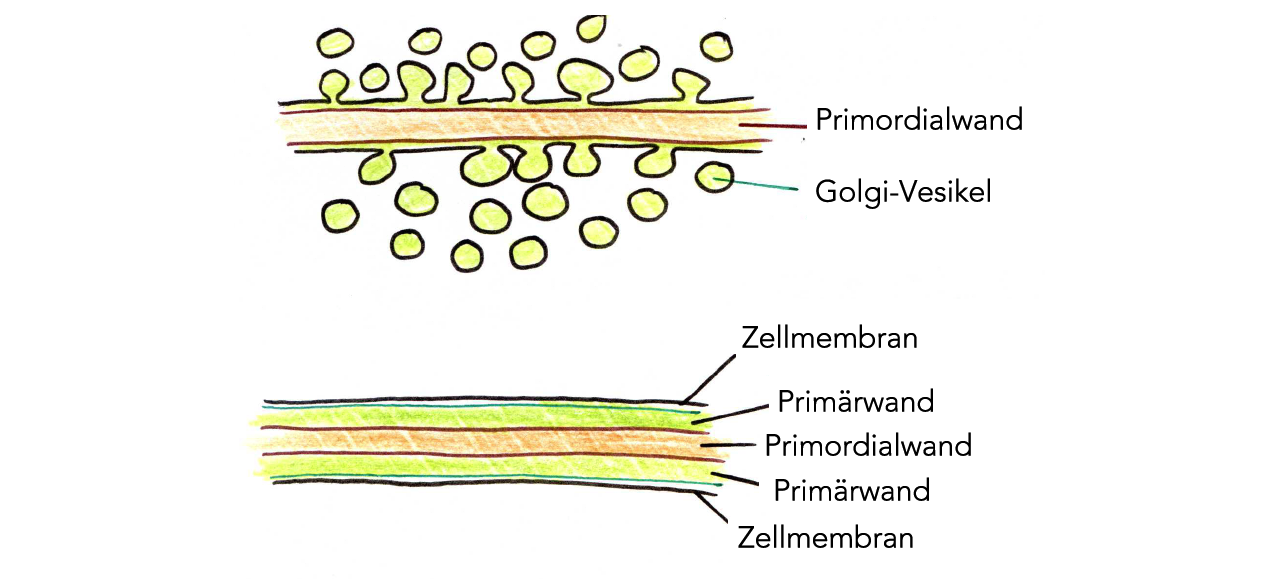
Auflagerung der Primärwand
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
In eine amorphe Matrix aus 50 bis 90% Pektinverbindungen sind zu ca. 5 bis 10% flexible und elastische Cellulosefibrillen eingelagert.
Bei Grünalgen enthalten die Fibrillen nur Cellulose, bei höheren Pflanzen kommen daneben Xylose, Arabinose und Mannose-Bausteine in den D-Glucoseketten vor, solche Polysaccharide bezeichnet man als Hemicellulosen.
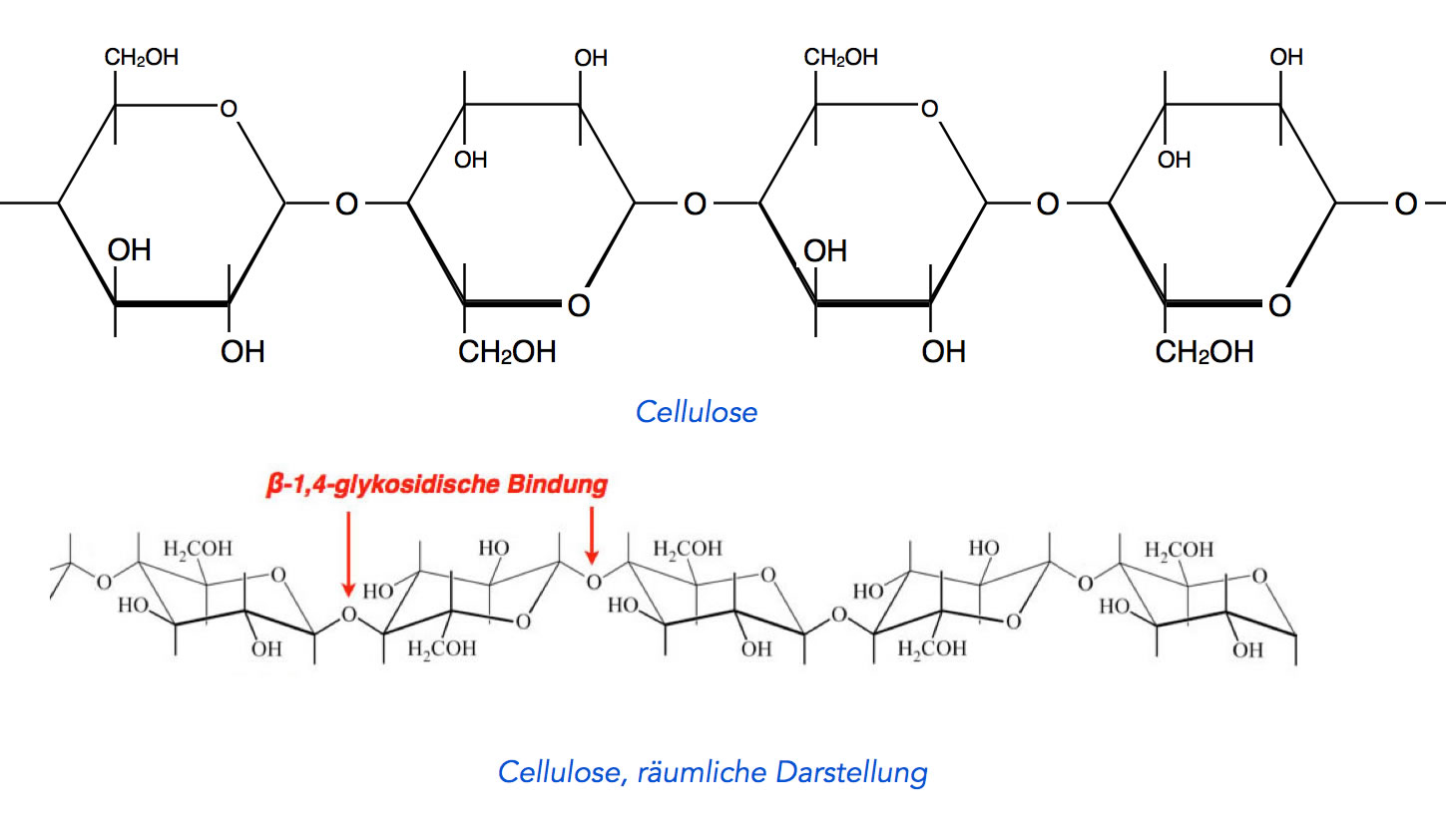
Cellulose
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Chemisch gesehen ist Cellulose ein Glucose-Polymer, in dem die einzelnen Glucose-Moleküle durch β-1,4-glycosidische Bindungen miteinander verbunden sind. Dabei polymerisieren ca. 14.000 Glucose-Moleküle zu einem Cellulose-Makromolekül.
Die Makromoleküle treten zu geordneten Einheiten, den Elementarfibrillen, zusammen. Die einzelnen Cellulose-Moleküle werden dabei durch Wasserstoffbrücken-Bindungen zusammengehalten. Eine Elementarfibrille enthält ca. 40 bis 100 Cellulose-Moleküle. Im Querschnitt sind die Elementarfibrillen ca. 3 bis 7 nm groß.
Die Elementarfibrillen sind ihrerseits zu sogenannten Mikrofibrillen organisiert. Eine Mikrofibrille enthält ca. 15 bis 20 Elementarfibrillen und somit 600 bis 2.000 Cellulose-Moleküle. Die Mikrofibrillen haben einen Durchmesser von ca. 10 bis 25 nm.
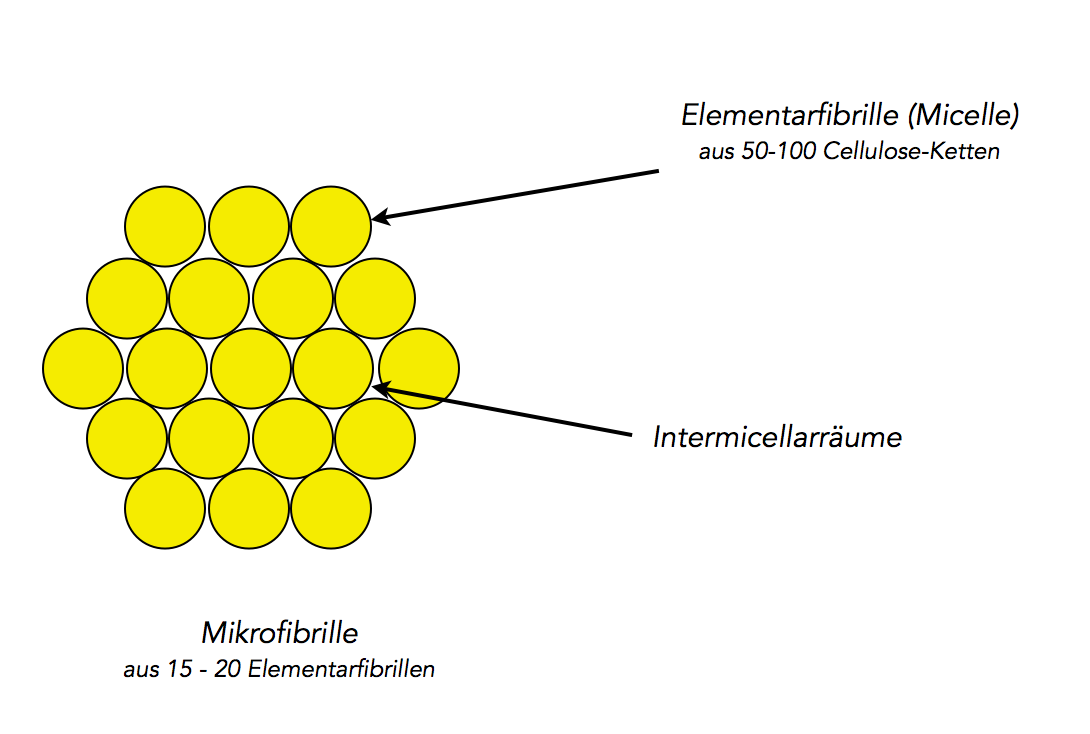
Mikrofibrillen bestehen aus 15 bis 20 Elementarfibrillen
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Die Intermicellarräume, also die Lücken zwischen den Elementarfibrillen in einer Mikrofibrille, spielen eine wichtige Rolle. Es können bestimmte Substanzen in sie eingelagert sein, welche die Eigenschaften der Zellwand verändern. Auch für einige mikroskopische Färbemethoden sind die Intermicellarräume wichtig, zum Beispiel bei der Chlor-Zink-Ion-Färbung.
Die Mikrofibrillen sind bereits im Elektronenmikroskop gut sichtbar. In der Primärwand sind die einzelnen Mikrofibrillen in der sogenannten Streuungstextur angeordnet, mit anderen Worten: Wahllos durcheinander:

Streuungstextur der Mikrofibrillen
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Bildung der Sekundärwand
Auf diese Primärwand, die nur ca. 10% Cellulose enthält, die in eine Pektinmatrix eingebettet ist, wird eine Sekundärwand aus 90% Cellulose aufgelagert. Auch das Material für diese Sekundärwand wird wieder von Golgi-Vesikeln der beiden Tochterzellen angeliefert.
Während die Mikrofibrillen der Primärwand in Streuungstextur angeordnet sind, liegen sie in der Sekundärwand in hochgeordneter Paralleltextur vor:
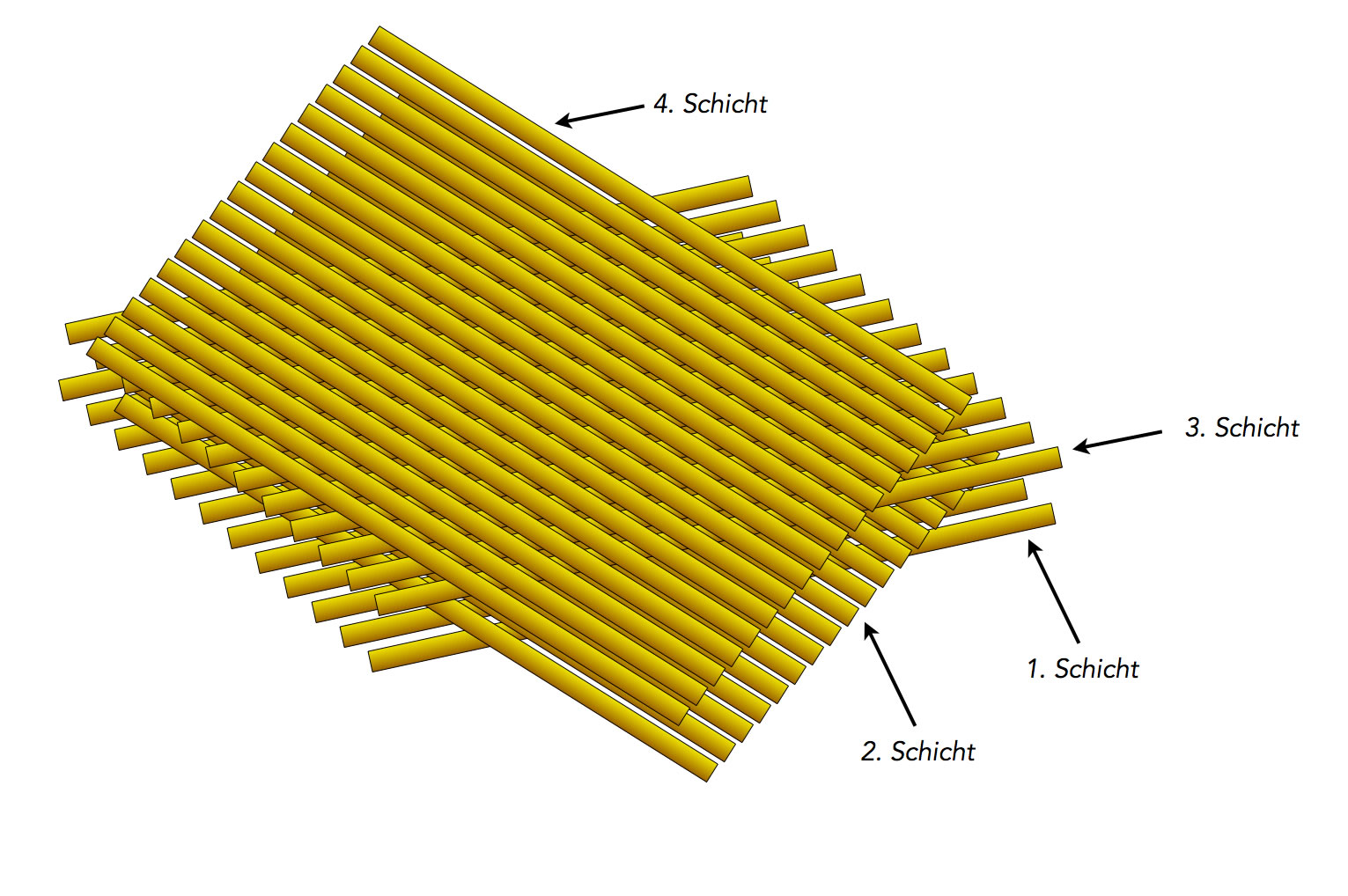
Eine Paralleltextur mit vier Schichten
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Dabei liegen mehrere Schichten übereinander. Die dritte Schicht entspricht in der Ausrichtung der Textur wieder der ersten Schicht, die vierte der zweiten und so weiter. Die einzelnen Cellulosefibrillen werden durch sogenannte Brückenpolysaccharide miteinander verbunden.
Tüpfel mit Plasmodesmen
Die Kommunikation erwachsener Pflanzenzellen bedarf wegen der existierenden Zellwände besonderer Vorrichtungen. In der Regel sind die Protoplasten benachbarter Zellen durch Plasmodesmen miteinander verbunden. Die einzelnen Plasmodesmen werden von speziellen ER-Ausläufern durchzogen.
Die Plasmodesmen durchziehen die Zellwand an bestimmten Stellen, den sogenannten Tüpfeln. Hier ist nur die Primordialwand oder Mittellamelle ausgebildet, Primärwand und Sekundärwand fehlen:
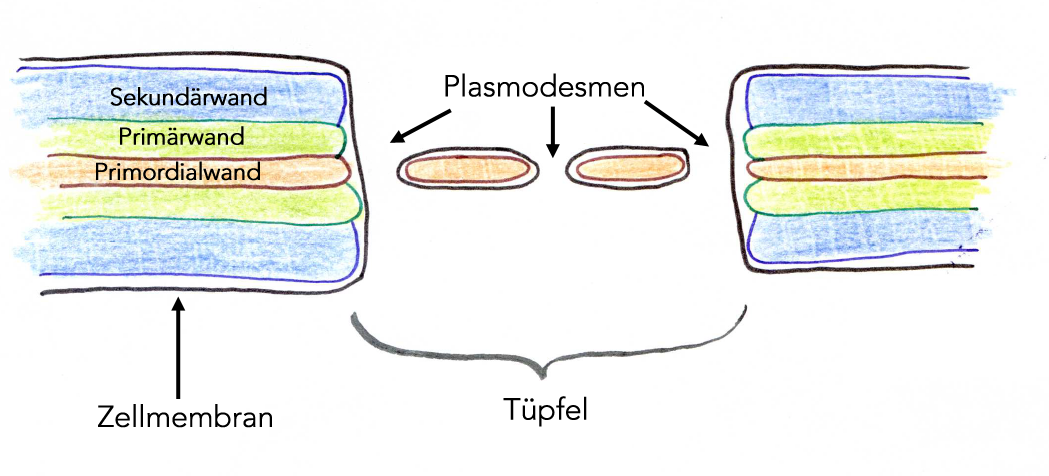
Aufbau eines Tüpfels im Querschnitt, es sind drei Plasmodesmen zu sehen.
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Hier eine weitere ältere Zeichnung:
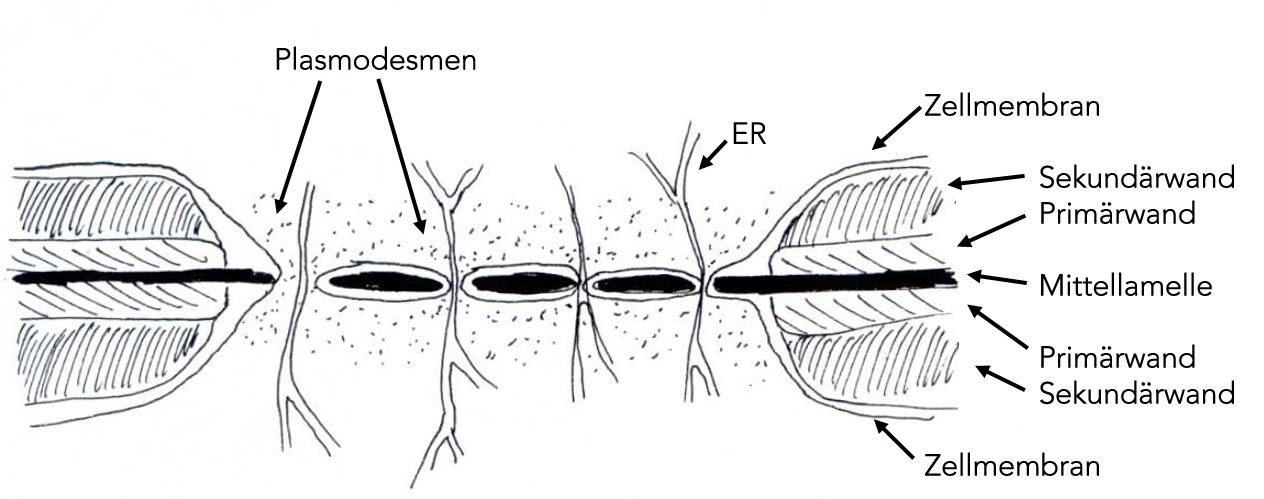
Eine Paralleltextur mit vier Schichten
Autor: Ulrich Helmich 1979/2021, Lizenz: Public domain
Ein Tüpfel enthält mehrere Plasmodesmen, und in einer Zellwand sind meistens mehrere bis viele Tüpfel eingelassen:
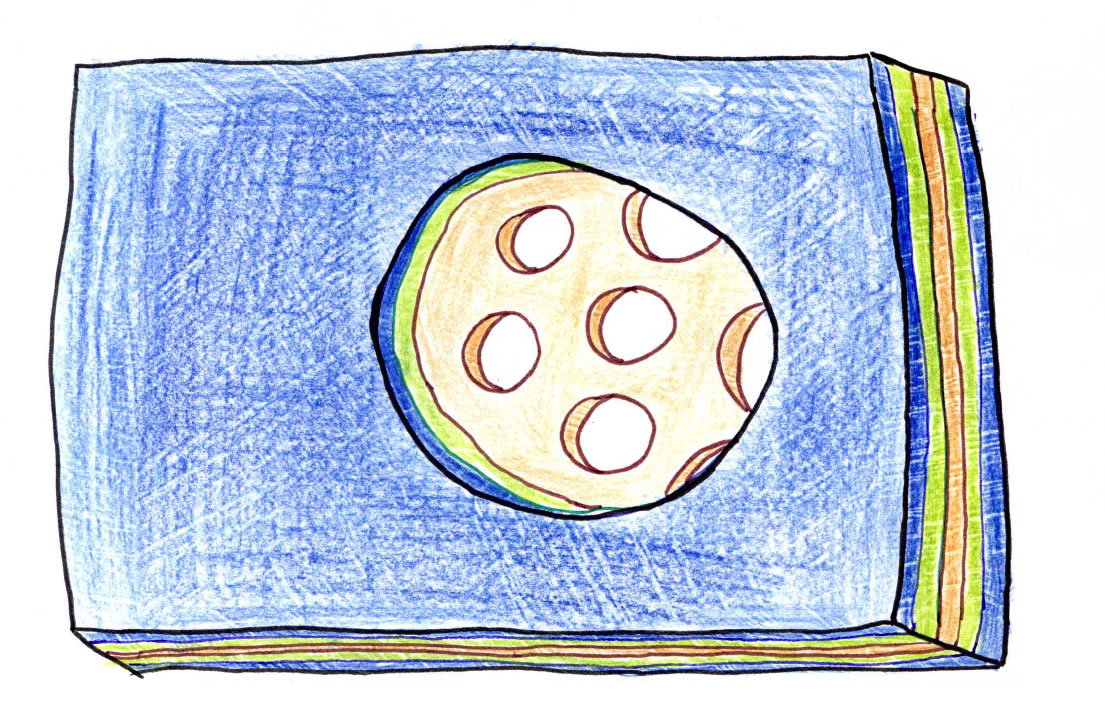
Ein Tüpfel
Autor: Ulrich Helmich 1978, Lizenz: Public domain
Die Primordialwand wird auch als Mittellamelle bezeichnet, weil sie genau in der Mitte der Zellwand liegt. Man beachte, dass die hier gezeichneten Zellwände stets zu zwei benachbarten Zellen gehören.
Anschließend kann noch eine innere Tertiärwand gebildet werden. Diese ist relativ dünn und besteht aus Cellulose und verschiedenen Matrixkomponenten.
Quellen:
- Kadereit , Körner, Nick, Sonnewald: Strasburger - Lehrbuch der Pflanzenwissenschaften, 38. Auflage, Springer Berlin Heidelberg 2021.
- Wikipedia, Artikel "Zellwand".
- Nultsch, Allgemeine Botanik, Stuttgart 1982.