Allgemeines
Eine einzelne Nervenzelle kann mit Hunderten oder sogar Tausenden anderer Nervenzelle verbunden sein. Mit anderen Worten, auf den Dendriten und auf dem Soma einer Nervenzelle können sich Hunderte oder Tausende von synaptischen Endigungen anderer Nervenzellen befinden. Jede einzelne synaptischen Endigung kann nun einen winzigen Bereich der postsynaptischen Membran depolarisieren oder hyperpolarisieren, je nachdem, ob es sich um eine erregende oder eine hemmende Synapse handelt.
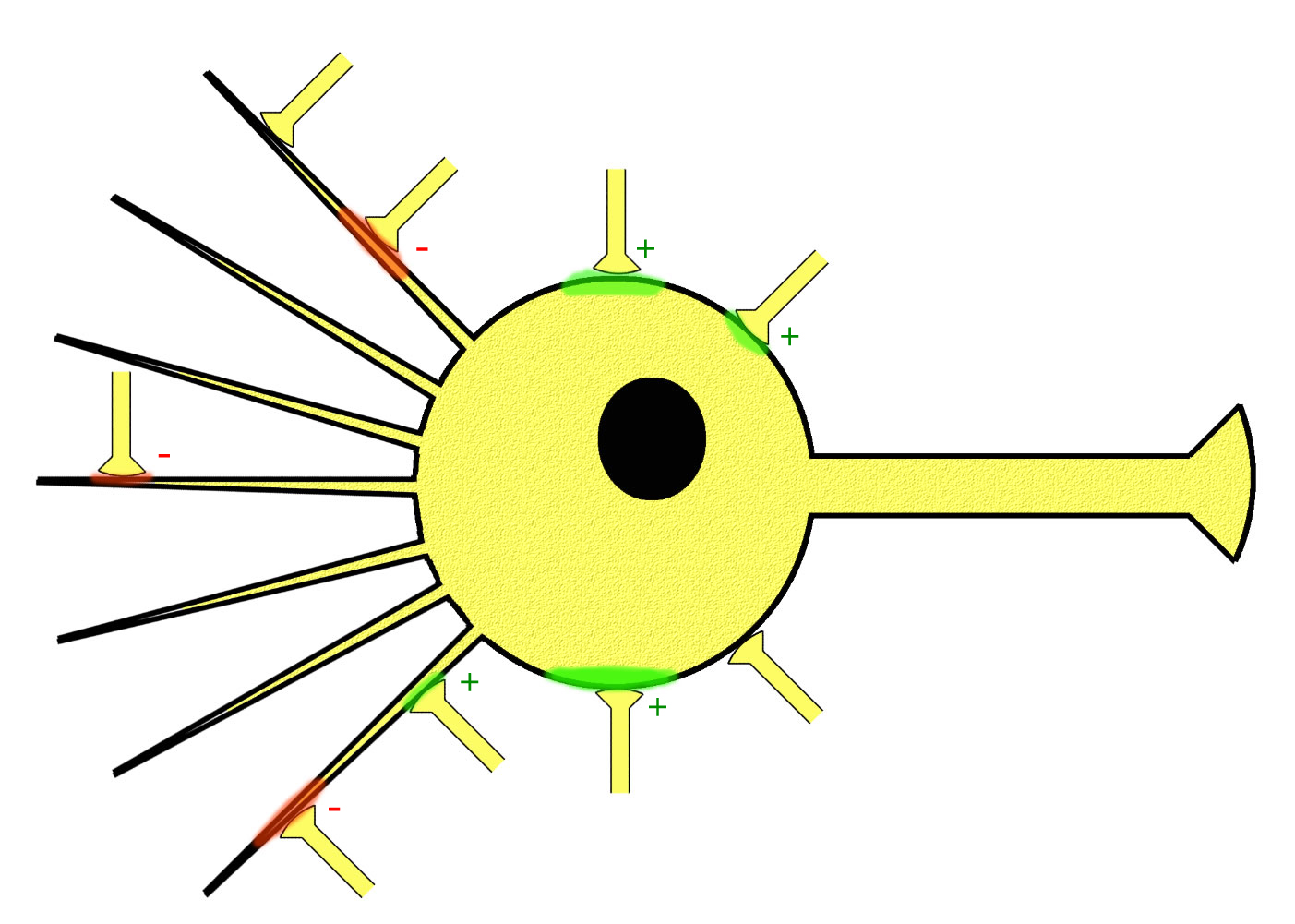
Verschaltung von Synapsen, Schema
Autor: Ulrich Helmich 2020, Lizenz: Public domain
Hier sieht man eine einfache Schemazeichnung einer Nervenzelle mit sieben Dendriten (in Wirklichkeit hat eine Nervenzelle sehr viel mehr dieser Dendriten) und neun Synapsen. Vier dieser neun Synapsen haben eine erregende Wirkung auf die Nervenzelle (grün, +), und drei Synapsen hemmen die Nervenzelle ( rot, -). Zwei Synapsen sind im Augenblick nicht aktiv.
Man kann gut sehen, dass einige Synapsen die Nervenzelle im Augenblick recht stark erregen bzw. hemmen, andere Synapsen dagegen nur recht schwach. Das Ausmaß der grünen bzw. roten Farbe soll die Intensität der Depolarisierung bzw. Hyperpolarisierung der postsynaptischen Membran aufzeigen.
Was passiert nun, wenn zwei, drei oder sogar sieben verschiedene Synapsen gleichzeitig aktiv sind, wie hier im Beispiel vier erregende und drei hemmende? Dazu siehe weiter beim Thema "räumliche Summation".
Was auf der schematischen Skizze nicht zu sehen ist: Ein und dieselbe synaptische Endigung kann auch für längere Zeit ihre Neurotransmitter ausschütten. Was passiert dann an der postsynaptischen Membran? Dazu siehe weiter beim Thema "zeitliche Summation".
Räumliche Summation
Von einer räumlichen Summation spricht man, wenn eine Nervenzelle gleichzeitig von mehreren präsynaptischen Zellen beeinflusst wird. Die Betonung liegt hierbei auf "gleichzeitig"!
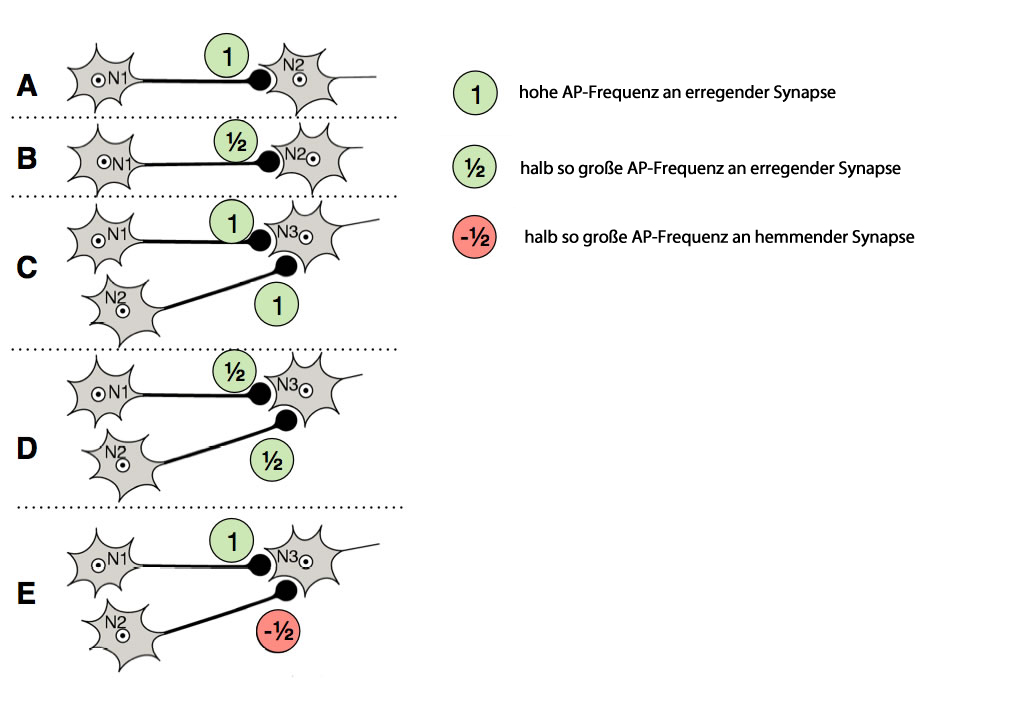
Beispiele für räumliche Summation
Autor: Ulrich Helmich 2020, Lizenz: Public domain
Auf dieser Schemazeichnung sieht man verschiedene Fallbeispiele für räumliche Summation. Ich möchte Ihnen diese fünf Beispiele jetzt näher erklären. Die klein gedruckten Abschnitte können Schüler(innen) der Sek. II gern überlesen, sie sind nur zur Vertiefung für Experten gedacht.
Fallbeispiel A
Die Empfängerzelle (rechts) ist nur mit einer Senderzelle verknüpft, die vielleicht mit einer Frequenz von 100/s Aktionspotenziale feuert. Die 100 APs/s an der erregenden Synapse depolarisieren die Membran der Empfängerzelle lokal. Es entsteht ein EPSP von vielleicht 40 mV, die postsynaptische Membran depolarisiert dann also von -70 mV auf -30 mV.
Am Axonhügel der Empfängerzelle ist dieses EPSP bereits stark abgeschwächt, es beträgt nur noch 10 mV, und mit einem resultierenden Membranpotenzial von -60 mV wird der Schwellenwert für die Ausbildung neuer Aktionspotenziale am Axonhügel noch nicht erreicht. Die Empfängerzelle bildet also keine Aktionspotenziale an ihrem Axon.
Nach neueren Erkenntnissen bewirkt bereits ein einziges Aktionspotenzial am synaptischen Endknöpfchen die Fusion von ca. 200 synaptischen Vesikeln mit der präsynaptischen Membran. Jedes Vesikel enthält über 1000 Neurotransmitter-Moleküle. Das EPSP, das durch ein Aktionspotenzial bewirkt wird, beträgt ungefähr 40 mV. Das heißt, das Membranpotenzial steigt von -70 auf -30 mV [1, S. 142].
Fallbeispiel B
Das gleiche Beispiel wie in A, allerdings können am Axon der Senderzelle nur halb so viele Aktionspotenziale pro Sekunde gemessen werden wie im Beispiel A, in unserem Fall also 50 APs/s. Das resultierende EPSP beträgt nur 20 mV, die Membran wird also von -70 auf -50 mV depolarisiert. Wenn die Depolarisierungswelle am Axonhügel ankommt, hat sie sich komplett abgeschwächt, das Membranpotenzial bleibt hier unverändert -70 mV.
Sitzt eine Synapse an einem Dendriten und schüttet das synaptische Endknöpfchen Neurotransmitter aus, dann wird die Membran des Dendriten an dieser Stelle depolarisiert (bei einer erregenden Synapse), es entsteht ein EPSP von vielleicht 40 mV, die Membran wird also von -70 mV auf -30 mV depolarisiert. Diese Depolarisierung wandert nun über Ausgleichsströmchen über die Membran des Dendriten in Richtung Soma. Dabei schwächt sich das EPSP aber ständig ab, und zwar exponentiell mit zunehmender Entfernung. Als Längskonstante bezeichnet man die Entfernung, an der die Depolarisation genau 37% des ursprünglichen Wertes beträgt. Der Wert dieser Längskonstante hängt hauptsächlich von zwei Faktoren ab, nämlich dem Längswiderstand des Dendriten und seinem Membranwiderstand [1, S. 143f]. Je größer der Längswiderstand, desto stärker wird das EPSP (oder IPSP) abgeschwächt. Je größer der Membranwiderstand, desto gering die Abschwächung durch Ionenströme quer durch die Membran.
Viele Dendriten von Gehirnneuronen enthalten spannungsgesteuerte Natrium- und Kaliumkanäle. Diese Kanäle öffnen sich, sobald das Membranpotenzial einen bestimmten Wert erreicht. Die dann ein-/ausströmenden Na+/K+-Ionen können das weitergeleitete EPSP/IPSP verstärken und so die Längskonstante erhöhen. Aktionspotenziale können an Dendriten jedoch nicht gebildet werden [1, S. 145].
Fallbeispiel C
Die Empfängerzelle wird gleichzeitig von zwei Senderzellen erregt. An jeder Synapse kommen 100 APs/s an, beide Stellen der postsynaptischen Membran werden auf -30 mV depolarisiert. Die beiden 40 mV-EPSPs bzw. Depolarisierungs-Wellen (elektrostatische Ausbreitung der Depolarisierung ähnlich wie die Wellen eines Steins im Wasser) summieren sich zu einem stärkeren EPSP von vielleicht 60 mV, das dann den Axonhügel erreicht und die Membran von -70 mV auf -10 mV depolarisiert. Damit wird der Schwellenwert für Aktionspotenziale schon deutlich überschritten. Am Axonhügel entstehen Aktionspotenziale. Die Frequenz dieser Aktionspotenzial hängt davon ab, wie stark der Schwellenwert am Axonhügel überschritten wird.
Fallbeispiel D
Die Empfängerzelle wird gleichzeitig von zwei Senderzellen erregt. An jeder Synapse kommen allerdings nur 50 AP/s an, was EPSPs von ca. 20 mV zur Folge hat und zu Depolarisierungen von -70 mV auf -50 mV führt.
Die beiden EPSPs addieren sich zu einem Summen-EPSP von vielleicht 30 mV, das dann in abgeschwächter Form den Axonhügel erreicht, so dass hier der Schwellenwert für Aktionspotenziale nicht überschritten wird.
Fallbeispiel E
Die Empfängerzelle wird gleichzeitig von zwei Senderzellen beeinflusst. Eine Senderzelle erregt die Empfängerzelle und erzeugt an der postsynaptischen Membran ein EPSP von vielleicht 40 mV, die andere Senderzelle hemmt die Empfängerzelle und erzeugt ein IPSP von vielleicht -20 mV. Allerdings ist die Hemmung schwächer als die Erregung, so dass sich in der Summe ein EPSP bildet von vielleicht 10 mV, das den Axonhügel aber höchstwahrscheinlich nicht erreicht, weil es sehr schwach ist. Sollte es dennoch den Axonhügel erreichen, wird das Membranpotenzial hier den Schwellenwert für Aktionspotenziale nicht überschreiten.
Die Neurotransmitter hemmender Synapsen setzen sich in Chlorid-Kanäle der postsynaptischen Membran und öffnen diese, so dass Chlorid-Ionen in die Zelle einströmen können. Nun liegt das Gleichgewichtspotenzial für Chlorid-Ionen bei -65 mV, also in der Nähe des Ruhepotenzials. Wenn also das Membranpotenzial der postsynaptischen Membran -65 mV beträgt oder noch negativer ist (-70 mV, -80 mV), dann hat das Öffnen der Chlorid-Kanäle keine besonderen Folgen für das Membranpotenzial, weil das Membranpotenzial ja schon ungefähr mit dem Chlorid-Potenzial identisch ist.
Interessanter ist die Frage, was passiert, wenn die postsynaptische Membran bereits durch erregende Synapsen depolarisiert ist, zum Beispiel auf -30 mV. Dann hat der Ausstrom der Chlorid-Ionen durchaus Folgen für das Membranpotenzial, es nimmt nämlich wieder ab in Richtung Gleichgewichtspotenzial der Chlorid-Ionen (-65 mV). Ein erzeugtes EPSP wird durch hemmende Synapsen also abgeschwächt.
Räumliche Lage der Synapsen
Bei der Summation der von den einzelnen Synapsen ausgelösten EPSPs und IPSPs spielt auch die räumliche Lage der jeweiligen Synapse eine wichtige Rolle. Je weiter die Synapse vom Axonhügel der Empfängerzelle entfernt ist, desto stärker schwächt sich das EPSP bzw. IPSP auf dem Weg zum Axonhügel ab, und desto geringer ist der Einfluss dieser Synapse auf das Zustandekommen von Aktionspotenzialen am Axonhügel.
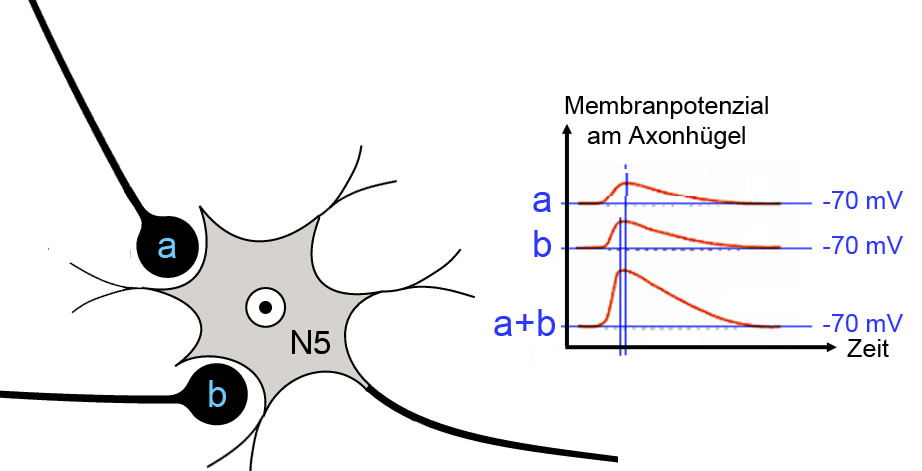
Einfluss der räumlichen Lage der Synapsen
Autor: Ulrich Helmich 2020, Lizenz: Public domain
Hier sehen Sie, wie sich die Lage der beiden erregenden Synapsen a und b auf das Membranpotenzial am Axonhügel auswirkt.
Die beiden Synapsen a und b haben nicht die gleiche Entfernung zum Axonhügel. Synapse a ist weiter entfernt als Synapse b.
- Wenn Synapse a 100 Neurotransmitter-Einheiten abgibt, steigt das Membranpotenzial am Axonhügel an, vielleicht von -70 auf -60 mV.
- Gibt Synapse b 100 Neurotransmitter-Einheiten ab, so wird ebenfalls ein EPSP erzeugt, allerdings kommt davon am Axonhügel etwas mehr an, und das EPSP erreicht den Axonhügel auch etwas eher als das EPSP der Synapse a. Das Membranpotenzial steigt am Axonhügel vielleicht von -70 auf -50 mV.
- Wenn beide Synapsen gleichzeitig Neurotransmitter abgeben, liegt eine räumliche Summation vor, und das Membranpotenzial am Axonhügel steigt auf vielleicht -40 mV. Das könnte schon für die Auslösung von Aktionspotenzialen am Axonhügel reichen, wenn der Schwellenwert entsprechend niedrig ist.
Räumliche Summation
Jede Nervenzelle wird gleichzeitig von vielen Synapsen erregt oder gehemmt. Bei der räumlichen Summation werden die von den verschiedenen Synapsen gebildeten EPSPs und IPSPs am Zellkörper verrechnet. Synapsen, die nahe am Axonhügel liegen, haben dabei einen größeren Einfluss als weit vom Axonhügel entfernte Synapsen. Überschreitet die Depolarisierung am Axonhügel einen Schwellenwert, so bilden sich dort neue Aktionspotenziale.
Inhibitorische Synapsen kommen auf den Dendriten eines Neurons vor, etwas häufiger auf dem Soma, und ganz besonders oft auf dem Axonhügel. Der Axonhügel ist eine strategisch wichtige Position für hemmende Synapsen. Eine geringe AP-Frequenz reicht hier schon aus, um die erregenden Impulse vieler anderer Synapsen zu hemmen [1, S. 148].
Zeitliche Summation
Die zeitliche Summation ist eigentlich recht einfach zu verstehen. Wenn ein Aktionspotenzial am synaptischen Endknöpfchen ankommt, wird eine bestimmte Menge von Neurotransmittern in den synaptischen Spalt ausgeschüttet. Diese Neurotransmitter setzen sich an entsprechende Rezeptoren der postsynaptischen Membran und führen zum Öffnen von Ionenkanälen; bei erregenden Synapsen sind dies Na+/K+-Kanäle.
Nach kurzer Zeit (Millisekunden) lösen sich diese Neurotransmitter aus den Rezeptoren und gelangen wieder in den synaptischen Spalt. Ein Teil der Neurotransmitter-Moleküle setzt sich erneut in die Rezeptoren (Ping-Pong-Prinzip), ein anderer Teil wird durch bestimmte Enzyme im synaptischen Spalt abgebaut oder von Gliazellen aufgenommen, so dass die Neurotransmitter unwirksam werden. Auf diese Weise dauert das erregende oder hemmende postsynaptische Potenzial (EPSP bzw. IPSP) nur eine kurze Zeit, in der Regel nur wenige Millisekunden.
Kommen in dieser Zeit aber weitere Aktionspotenziale am gleichen synaptischen Endknöpfchen an, so werden weitere Neurotransmitter in den synaptischen Spalt freigesetzt. Die Rezeptoren der postsynaptischen Membran werden also weiterhin durch Neurotransmitter besetzt, und bei einer erregenden Synapse fließen weitere Natrium-Ionen in die postsynaptische Zelle ein. Die Depolarisierung der postsynaptischen Membran hält also nicht nur an, sondern wird sogar verstärkt.
Im Grunde ist das schon alles, was man zur zeitlichen Summation wissen muss. Nichtsdestotrotz möchte ich Ihnen hier mit einer kleinen Zeichnung "auf die Sprünge" helfen, damit Sie die zeitliche Summation noch besser verstehen. Ähnliche Skizzen hat man auch schon im Biologie-Abitur (schriftlich / mündlich) gesehen.
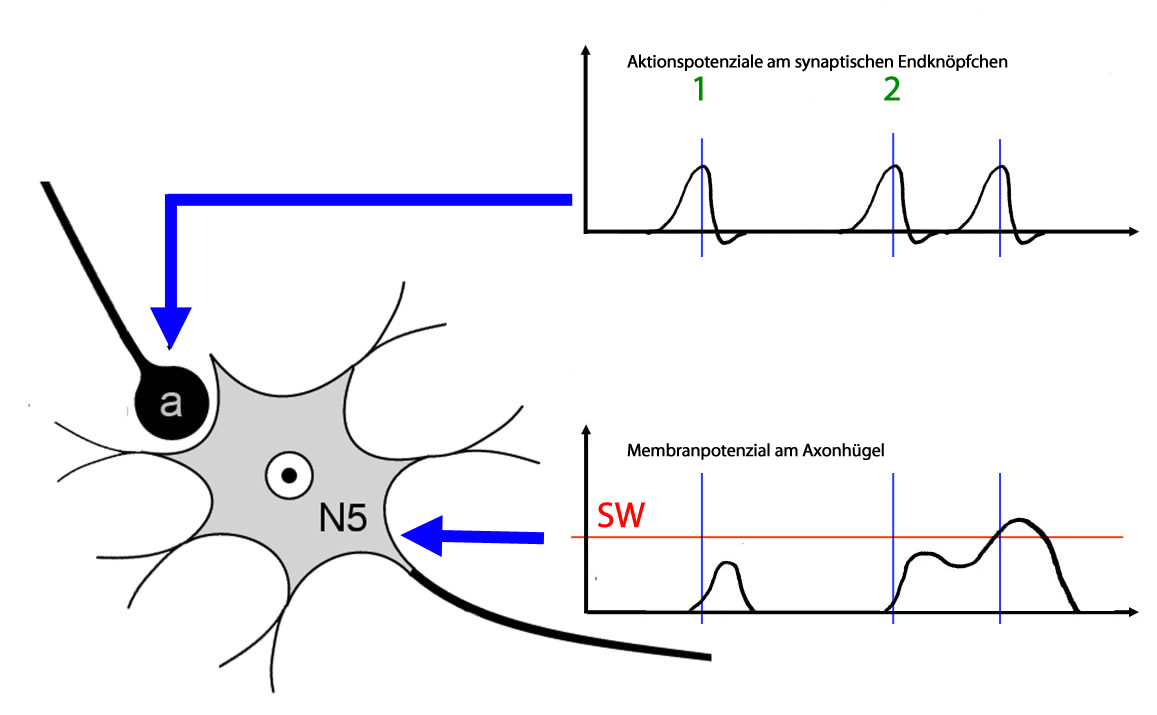
Zeitliche Summation
Autor: Ulrich Helmich 2020, Lizenz: Public domain
Hier ist nur eine Synapse zu sehen. Zum Zeitpunkt 1 wird diese Synapse nur einmal aktiviert, das Messgerät zeichnet nur ein Aktionspotenzial auf, und es wird nur eine "Portion" Neurotransmitter ausgeschüttet.
Am Axonhügel ein kleines EPSP, das von dem zweiten Messgräte (unteres Diagramm) aufgezeichnet wird.
Zum Zeitpunkt 2 kommen zwei Aktionspotenziale am Endköpfchen an (oberes Diagramm). Es werden zwei "Portionen" Neurotransmitter in den synaptischen Spalt abgegeben, und die beiden ausgelösten EPSPs summieren sich zu einem größeren Summenpotenzial, das am Axonhügel den Schwellenwert (SW) für Aktionspotenziale überschreitet. Die dünnen blauen Linien in den beiden Diagrammen sollen verdeutlichen, dass es ein paar Millisekunden dauert, bis ein EPSP am Axonhügel ankommt.
Quellen, die über allgemeines Schulbuchwissen hinausgehen:
- Bear, Connors, Paradiso: Neurowissenschaften, Springer-Verlag 2018