Das Schulbuch-Modell
Auf dieser Seite wollen wir zunächst das einfache "Schulbuch-Modell" des lac-Operons besprechen, also so, wie es in fast jedem Oberstufen-Lehrbuch der Biologie dargestellt ist. Auf einer Vertiefungsseite gehen wir dann auf die neueren Erkenntnisse zum lac-Operon ein; immerhin ist das Operon-Modell schon in den 60er Jahren des letzten Jahrhunderts entstanden, und in der Zwischenzeit hat sich einiges getan.
Aufbau eines prokaryotischen Operons
Betrachten wir dazu das folgende Bild:
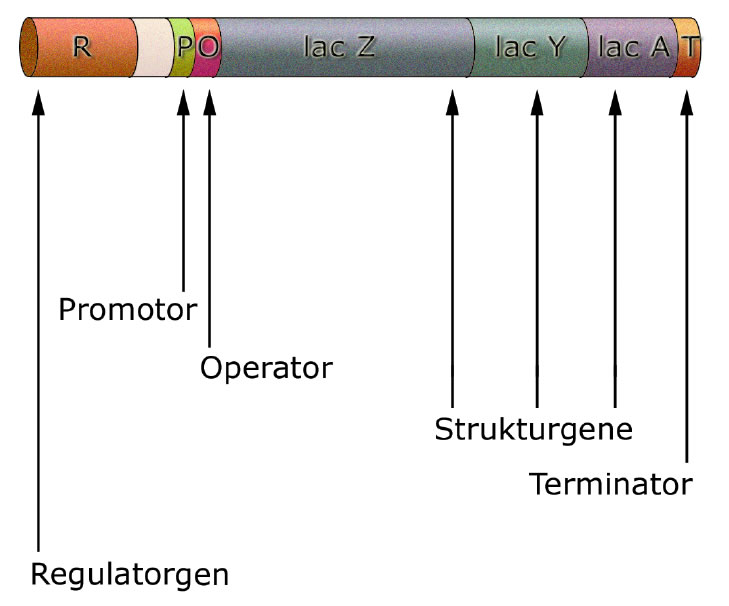
Aufbau des lac-Operons
Autor: Ulrich Helmich 2013, Lizenz: Public domain.
Hier sehen wir den Aufbau des lac-Operons. Die Region R ganz links wollen wir zunächst einmal ignorieren, wir kommen im nächsten Abschnitt auf sie zurück.
Das eigentliche lac-Operon beginnt mit dem Promotor. Der Promotor ist die Ansatzstelle für die RNA-Polymerase, hier beginnt das Enzym mit der Transkription der lac-Gene.
Für alle, die sich nicht mehr ganz sicher sind, wie die Transkription abläuft...
Es folgt der Operator, eine Steuereinheit, die darüber entscheidet, ob die drei Strukturgene transkribiert werden oder nicht.
Dann kommen die drei Strukturgene lac Z, lac Y und lac A. Diese drei Gene enthalten den Code für Enzyme, die für den Abbau der Lactose verantwortlich sind.
Das lac-Z-Gen ist für das Enzym ß-Galactosidase verantwortlich, das Lactose in seine beiden Bestandteile Glucose und Galactose spaltet. Die Glucose kann dann ganz normal über die Glycolyse abgebaut werden.
Das lac-Y-Gen enthält die Bauanleitung für eine Permease. Das ist ein Enzym, welches sich in die Zellmembran der Bakterienzelle setzt und für den Transport der Lactose in die Zelle hinein verantwortlich ist.
Das lac-A-Gen schließlich codiert eine Transacetylase. Die Funktion dieses Enzyms ist zur Zeit (2022) noch nicht vollständig bekannt. Sicher ist nur, dass das Enzym eine Acetylgruppe von Acetyl-CoA auf die Lactose überträgt. Eine Autorengruppe vermutet, dass dieses Enzym Verbindungen "entgiftet", die von der Permease mit in die Zelle transportiert werden [1].
Am Terminator stoppt die RNA-Polymerase mit der Transkription.
Ein prokaryotisches Operon besteht aus einem Promotor, einem Operator, einer Reihe von funktionell zusammengehörigen Strukturgenen und einem Terminator.
Das Operon kann reguliert werden
Kommen wir nun zur Rolle des Operators und des Regulatorgens.
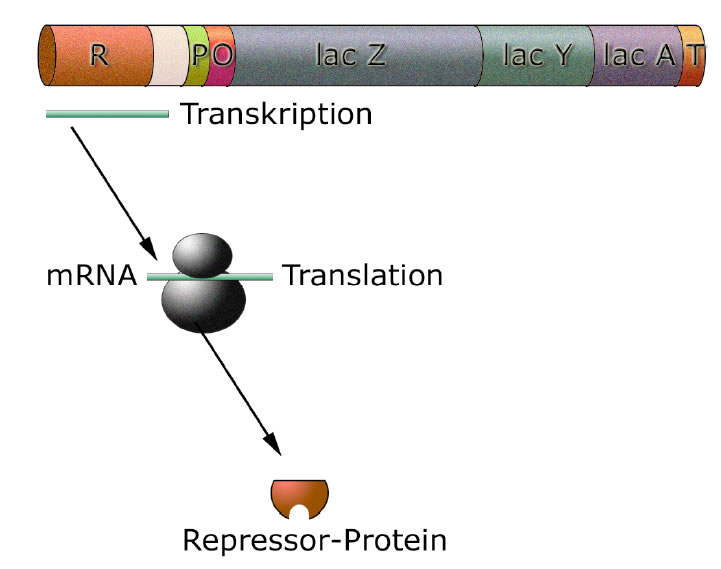
Das Regulatorgen ist für die Expression eines Repressorproteins zuständig
Das Regulatorgen R codiert die Bauanleitung für ein Repressor-Protein. Die Abbildung zeigt die Bildung dieses Repressor-Proteins. Das Regulatorgen wird transkribiert, die mRNA translatiert, und das gebildete Protein muss dann noch prozessiert werden (was hier nicht zu sehen ist).
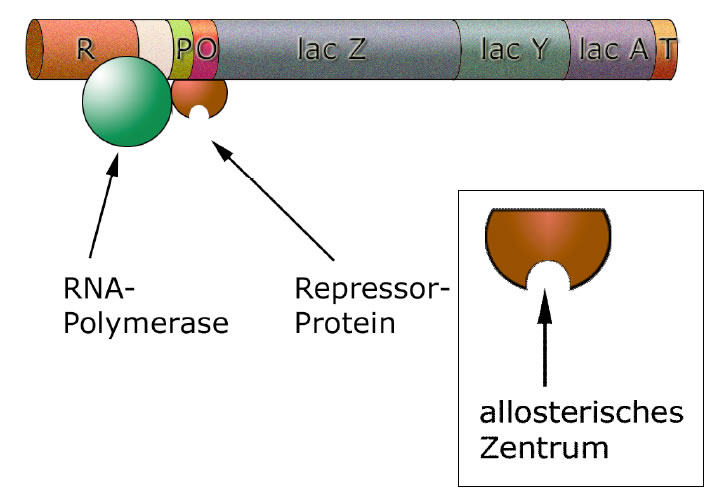
Der Repressor blockiert die RNA-Polymerase
Das Repressor-Protein (oder kurz: der Repressor) setzt sich nach dem Schlüssel-Schloss-Prinzip an den Operator des lac-Operons und blockiert dadurch die RNA-Polymerase. Wenn der Repressor am Operator sitzt, kommt die große Polymerase nicht mehr an dem Repressor vorbei, somit wird die Transkription abgebrochen.
Das Repressorprotein hat ein allosterisches Zentrum, das auf der Zeichnung auch deutlich zu sehen ist (übertrieben groß).
Jetzt überlegen wir einmal: Wenn genügend Lactose im Außenmedium vorhanden ist, wäre die Bakterienzelle ja schön dumm, wenn sie diese Nahrungsquelle nicht nutzen würde. Dazu müssen die Lactose-Gene aber transkribiert werden. Der Repressor muss also "aus dem Weg geräumt" werden, wenn genug Lactose vorhanden ist.
Können Sie sich nun vorstellen, welche Aufgabe das allosterische Zentrum des Repressors hat? Wenn Sie noch nicht von allein auf die Lösung gekommen sind, schauen Sie sich doch einfach das nächste Bild an:
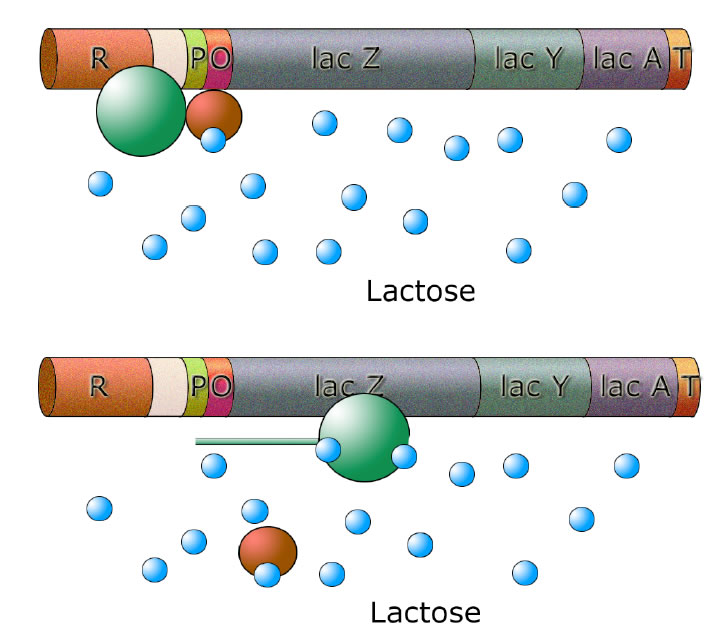
Lactose setzt sich in das allosterische Zentrum; der Repressor gibt den Weg für die Polymerase frei
Das allosterische Zentrum des Repressors kann ein Lactose-Molekül aufnehmen. Dadurch verändert der Repressor seine Struktur (Konformations-Änderung), und plötzlich passt er nicht mehr an den Operator (Schlüssel-Schloss-Prinzip: Der Schlüssel wurde verbogen und passt nicht mehr in das Schloss...).
Der Repressor löst sich jetzt von dem Operator und gibt so den Weg für die RNA-Polymerase frei. Es wird mRNA gebildet für die Produktion der drei Enzyme, die Lactose abbauen und energetisch verwerten.
Und wie geht es weiter?
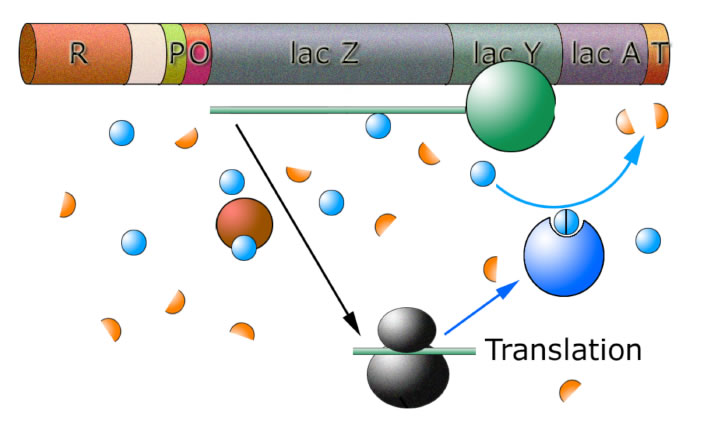
Abbau der Lactose durch die exprimierten Enzyme
Durch die Aktivität der Lactose-abbauenden Enzyme sinkt die Lactose-Konzentration in der Zelle. Wenn sich ein Lactose-Molekül in das allosterische Zentrum eines Repressors sitzt, so ist diese Bindung nicht von Dauer. Die Lactose dockt nur locker an, ein paar Millisekunden später kann sich das Molekül auch wieder aus dem Repressor lösen, und wieder ein paar Millisekunden später kommt ein anderes Lactose-Molekül und setzt sich wieder in das allosterische Zentrum.
An dieser Stelle ist also dynamisches Denken gefragt, nicht statisches. Wir müssen mit Wahrscheinlichkeiten rechnen. Ein Teil der 10 bis 20 lac-Repressor-Moleküle einer E. coli-Zelle ist zu einem bestimmten Zeitpunkt mit Lactose-Molekülen besetzt, ein anderer Teil ist gerade nicht besetzt. Die Wahrscheinlichkeit, einen mit Lactose besetzten Repressor anzutreffen, wird geringer, wenn die Lactose-Konzentration sinkt. Je mehr Lactose also abgebaut wird, desto unwahrscheinlicher ist es, dass ein Repressor Lactose in seinem allosterischen Zentrum sitzen hat.
Mit sinkender Lactose-Konzentration steigt also die Wahrscheinlichkeit, dass die Operator-Region durch ein Repressor-Molekül blockiert wird, und die Transkriptionsrate sinkt gegen null. Dies ist ja auch sinnvoll, wenn keine Lactose mehr vorhanden ist.
Übrigens wird diese Art und Weise der Genregulation auch Substratinduktion genannt, weil das abzubauende Substrat - in diesem Fall die Lactose - seinen eigenen Abbau induziert (auslöst).
Substratinduktion
Ein Substrat wie Lactose, Glucose etc. löst seinen eigenen Abbau durch Enzyme aus.
Ist die Substratkonzentration gering, so sind die Strukturgene dieser Enzyme durch ein Repressor-Protein blockiert, das am Operator angedockt ist (Schlüssel-Schloss-Prinzip). Die RNA-Polymerase wird vom Repressor abgeblockt und kann die Strukturgene nicht transkribieren.
Ist die Substratkonzentration dagegen hoch, so setzen sich Substrat-Moleküle (oder andere Effektor-Moleküle) in die allosterischen Zentren der Repressoren, diese ändern ihre Konformation, lösen sich von den Operatoren und machen den Weg für die RNA-Polymerase frei. Die Strukturgene werden transkribiert und translatiert, es werden Enzyme gebildet, die das Substrat abbauen. Dadurch sinkt die Substratkonzentration, die Repressor-Proteine nehmen wieder die ursprüngliche Konformation an und blockieren erneut die DNA-Polymerase.
Quellen, die über allgemeines Schulbuchwissen hinausgehen:
- Wang, Olsen, Roderick: "Structure of the lac Operon Galactoside Acetyltransferase", Cell Vol. 10, Issue 4, p. 581-588, 2002.