Erleichterte Diffusion durch Transportproteine
Carrierproteine hatten wir bereits auf der entsprechenden Seite kennen gelernt. Carrierproteine ermöglichen ähnlich wie Kanalproteine eine erleichterte Diffusion. Normalerweise können hydrophile Teilchen die Lipid-Doppelschicht nur sehr schwer passieren, nach einiger Zeit schaffen sie es jedoch. Kanalproteine und Carrierproteine erleichtern diese Diffusion aber enorm, weil sie eine hydrophile Pore bzw. einen Klapptür-Mechanismus zur Verfügung stellen. Allerdings kann dieser Transport nicht mit der gleichen hohen Geschwindigkeit erfolgen wie eine freie Diffusion ohne "störende" Membran.
Aktiver Transport
In vielen Situationen müssen Solute (das sind die zu transportierenden Stoffe, die im Gegensatz zu einem Substrat durch den Transportvorgang nicht chemisch verändert werden) gegen ein bestehendes Konzentrationsgefälle in die Zelle oder in ein bestimmtes Organell hinein transportiert werden (oder heraus).
Ein Beispiel für einen solchen Vorgang ist der Glucose-Transport in bestimmte Zellen. Obwohl die Zellen bereits viel von dem Nährstoff enthalten, sammeln sie weitere Glucose aus dem Außenmedium an. Möglich wird dies durch einen aktiven Transport.
Ein weiteres Beispiel ist die Lichtreaktion der Photosynthese bei den grünen Pflanzen. Damit die Lichtenergie in chemische Energie umgewandelt werden kann, ist die Erzeugung eines Protonengradienten quer über die Membran der Chloroplasten-Thylakoide notwendig. Dieser Protonengradient entsteht nicht von selbst, sondern muss unter Energieaufwand erzeugt werden.
Ein drittes Beispiel sind die Vorgänge bei der Bildung von Aktionspotenzialen in der Nervenzelle. Nach einem Aktionspotenzial befinden sich in der Zelle zu viele Natrium-Ionen. Diese müssen wieder aus der Zelle heraus transportiert werden. Allerdings ist die Konzentration der Natrium-Ionen außerhalb der Zelle sehr hoch, so dass ein Transport gegen das Konzentrationsgefälle notwendig wird.
Genau so wenig, wie Wasser von alleine nach oben fließt, kann man keinen Stoff gegen sein Konzentrationsgefälle transportieren, ohne dabei Arbeit zu leisten, die wiederum Energie erfordert (Energie-Umwandlung; ein "Verbrauch" von Energie ist nicht möglich, sagen die Physiker).
Es gibt nun im Prinzip drei Möglichkeiten, wie eine Zelle einen solchen aktiven Transport ( = Transport gegen ein Konzentrationsgefälle) durchführen kann.
1. Unter ATP-Verbrauch
Viele Carrierproteine sind in der Lage, die universelle Energiewährung ATP (Adenosintriphosphat) in ADP und anorganisches Phosphat Pi zu spalten. Bei dieser Hydrolyse wird sehr viel chemische Energie freigesetzt, die dann genutzt werden kann, um mechanische Arbeit zu leisten, wie es das "Bergauf"-Pumpen von Teilchen ja darstellt. Tatsächlich wird der aktive Transport gegen ein Konzentrationsgefälle oft auch als Bergauf-Transport oder Bergauf-Diffusion bezeichnet.
➥Arbeitsweise der Natrium-Kalium-Pumpe
Auf meinen Neurobiologie-Seiten für die Qualifikationsphase finden Sie eine sehr schönes Beispiel, das zeigt, wie ein solcher aktiver Transport unter ATP-Verbrauch arbeitet.
Diese Art des aktiven Transports wird auch als direkter aktiver Transport bezeichnet. Die Energiequelle, nämlich ATP, ist direkt an den Transportvorgang gekoppelt.
2. Durch gekoppelten Transport
Ein Konzentrationsgefälle der Teilchenart A ist in der Lage, Arbeit zu leisten. Staut man Wasser in einem Staubecken und lässt es dann bergab fließen, kann man damit Pumpen oder Turbinen antreiben. Lässt die Zelle einen Stoff A mit dem Konzentrationsgefälle durch ein Carrierprotein die Membran passieren, so kann auch hier "nebenbei" Arbeit geleistet werden.
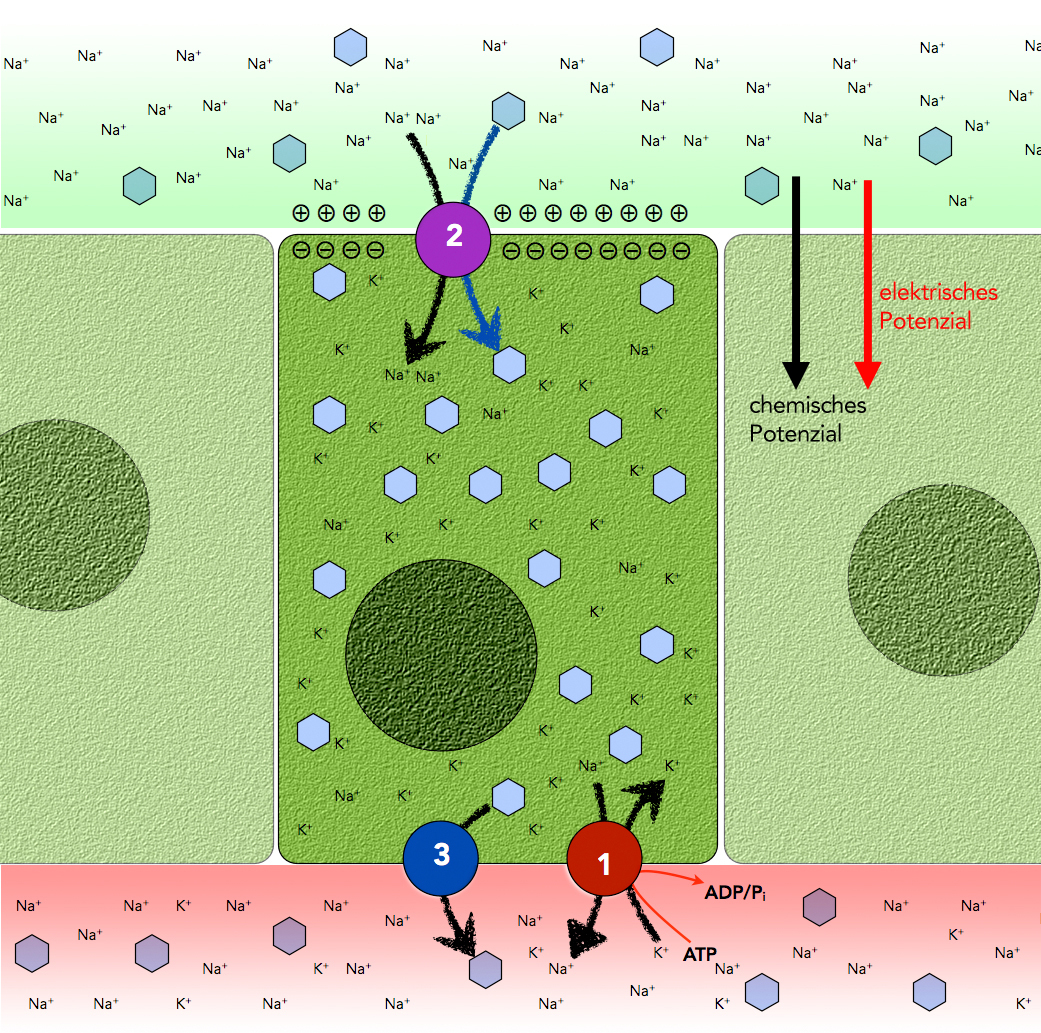
In dem Konzentrationsgradienten des Stoffes A ist ja viel Energie gespeichert, man spricht hier auch von einem chemischen Potenzial. Oft wird dieses chemische Potenzial auch noch von einem elektrischen Potenzial unterstützt. In Tierzellen wird meistens ein Na+-Gradient als Energiequelle genutzt. Außerhalb der Zelle herrscht eine hohe Na+-Konzentration, innerhalb der Zelle sorgt die Natrium-Kalium-Pumpe für eine dauerhaft niedrige Na+-Konzentration. Da die Innenseite der Membran negativ geladen ist, werden einströmende Na+-Ionen zusätzlich elektrisch angezogen. In diesem elektrochemischen Potenzial (Konzentrationsgradient + Membranspannung) steckt viel Energie, die benutzt werden kann, um Arbeit zu leisten. Diese Arbeit besteht bei dem gekoppelten Transport darin, einen anderen Stoff B gegen sein Konzentrationsgefälle zu transportieren.
Natürlich muss dafür die beim A-Transport freigesetzte Energie größer sein als die zum B-Transport erforderliche Energie, weil ja keine Energieumwandlung verlustfrei geschieht. Der Stoff B kann bei diesem gekoppelten Transport entweder in der gleichen Richtung erfolgen wie der Transport von A (Symport), oder in der entgegengesetzten Richtung (Antiport). In beiden Fällen handelt es sich um einen aktiven Transport von B. Allerdings spricht man hier von einem sekundären aktiven Transport. ATP ist ja nur indirekt für den B-Transport verantwortlich, indem es für den Aufbau des A-Gradienten sorgt.
2.1 Antiport
Ein Antiport
Autor: Ulrich Helmich 2021, Lizenz: Public domain
In dieser vereinfachten Darstellung sieht man zwei Teilchensorten, die durch rote Kugeln und grüne Dreiecke symbolisiert werden. Die grünen Dreiecke diffundieren passiv mit ihrem Konzentrationsgradienten in die Zelle. Die hierbei freigesetzte Energie wird für den aktiven Transport der roten Kugeln genutzt. Da die roten Kugeln in Gegenrichtung zu den grünen Dreiecken transportiert werden, spricht man hier auch von einem Gegentransport oder Antiport.
2.2 Symport
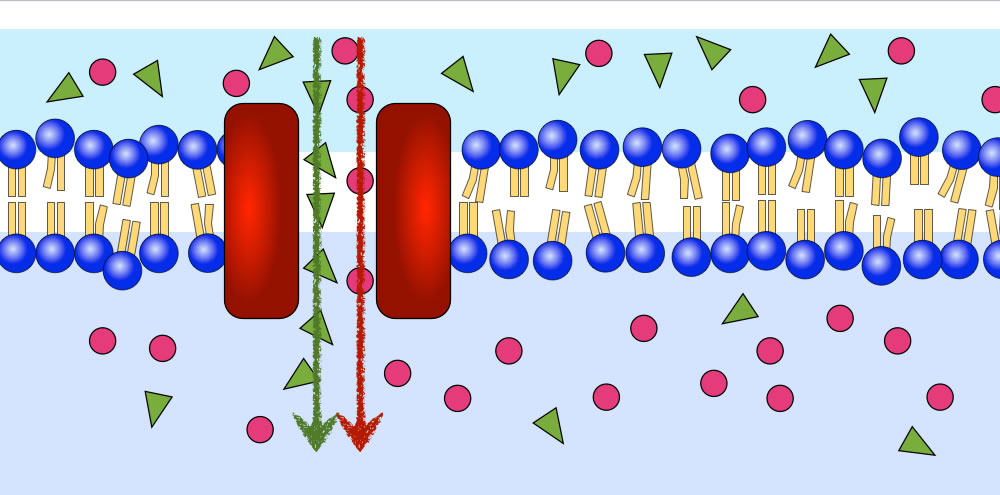
Ein Symport
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Der gekoppelte Transport der B-Teilchen kann auch in der gleichen Richtung stattfinden wie die passive Diffusion der A-Teilchen. In diesem Fall spricht man von einem Symport.
Anwendung: Der Na+-Glucose-Transporter
Wir besprechen nun als Nächstes ein lehrreiches Beispiel, in dem alle bisher besprochenen Transportarten vereinigt sind: Die Aufnahme von Glucose in die Epithelzellen des Dünndarms. Dieses Beispiel ist so gut und wichtig, dass es sogar Einzug in gute Oberstufen-Lehrbücher gefunden hat, zum Beispiel den aktuellen Linder.
ATP-getriebene Ionenpumpe, Symport und Antiport im Dünndarm
Bei der Verdauung von Kohlenhydraten im Dünndarm entsteht durch die Einwirkung zahlreicher Enzyme Glucose als Endprodukt. Diese Glucose muss jetzt vom Dünndarm in das Blut gelangen. Zunächst muss die Glucose aber in die Epithelzellen transportiert werden, die den Dünndarm auskleiden. Erst von da aus gelangt die Glucose schließlich in das Blut.
Glucose-Transport im Dünndarm
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Der Transport der Glucose vom Dünndarm in die Epithelzellen muss aktiv erfolgen, da in den Epithelzellen bereits eine hohe Glucose-Konzentration herrscht.
- Eine ATP-getriebene Natrium-Kalium-Pumpe betreibt einen Antiport: Kalium-Ionen werden gegen das K+-Gefälle in die Epithelzelle hinein transportiert, Natrium-Ionen werden im Gegenzug gegen das Na+-Gefälle aus der Zelle heraus transportiert, so dass dort stets eine sehr geringe Na+-Konzentration herrscht.
- Diese geringe Na+-Konzentration wird auch dringend benötigt für den nächsten Schritt. Aus dem Darmlumen diffundieren nun Na+-Ionen mit dem Konzentrations- und mit dem Ladungsgefälle (Innenseite negativ!) in die Epithelzelle hinein. Diese passive Transport ist gekoppelt mit einem Glucose-Bergauftransport in gleicher Richtung. Mit jeweils zwei Na+-Ionen wird ein Glucose-Molekül in die Zelle transportiert.
- Der Transport der Glucose aus der Epithelzelle in das Blut geschieht nun rein passiv, in Richtung des Konzentrationsgradienten. Es handelt sich um einen typischen Uniport.
Letztendlich ist also auch der aktive Transport der Glucose-Moleküle in die Epithelzellen hinein ein ATP-getriebener Transport, aber kein direkter, sondern ein indirekter über den Umweg des Na+-getriebenen Symporters. Daher spricht man hier auch von einem sekundären aktiven Transport. Neben Glucose kann übrigens auch das ähnlich aussehende Galactose-Molekül auf diese Weise transportiert werden [3].
Auf dieser Lexikonseite findet sich die gleiche Abbildung wie hier, allerdings wird zusätzlich auf den genauen Mechanismus des Transports eingegangen.
Wer sich für das Fach Ernährungslehre interessiert, kann auf dieser Seite alles über die Verdauung der Kohlenhydrate nachlesen.
Diese Art des gekoppelten Transports (Symport, Antiport) ist nicht auf Kohlenhydrate wie Glucose oder Galactose beschränkt. Auch viele Aminosäuren werden auf diese Art und Weise in die Zelle transportiert.
3. Lichtgetriebene Pumpen
Alle Zellen, auch prokaryotische, benötigen Energie für alle möglichen Zwecke. Die universelle Energiewährung der Zellen ist das ATP (Adenosintriphosphat). Um ATP herzustellen, lassen die meisten Zellen eine umgekehrte Protonenpumpe arbeiten.
Normale Protonenpumpe:
Ein Carrier transportiert Protonen gegen den H+-Gradienten bergauf durch eine Membran und verbraucht dabei ATP.
Umgekehrte Protonenpumpe:
Alles läuft rückwärts ab: Die Protonen fließen mit dem Konzentrationsgradienten durch den Carrier und erzeugen dabei ATP. Ein solches Carrierprotein wird dann aber nicht mehr als Carrier bezeichnet, sondern als ATPase, also als Enzym, das ATP herstellt.
Damit eine ATPase arbeiten kann, muss also zuvor ein Protonengradient erzeugt werden. Bei photosynthetisch aktiven Bakterien und den grünen Pflanzen wird dieser Protonengradient mit Hilfe von Lichtenergie aufgebaut. Wie das alles genau abläuft, habe ich auf meinen Seiten zur Photosynthese erläutert. Hier nur eine Kurzfassung: Die Lichtenergie treibt chemische Vorgänge an, die auf der einen Seite der Thylakoid-Membran Protonen freisetzen, während auf der anderen Seite der Thylakoid-Membran Protonen verbraucht werden, indem sie zum Beispiel in Transport-Moleküle eingebaut werden. Es müssen also gar keine Protonen quer durch die Membran transportiert werden, um einen Protonengradienten zu erzeugen.
Auf diese Art der Erzeugung von Protonengradienten kommen wir auch im folgenden Abschnitt zu sprechen.
4. Durch Redoxvorgänge angetriebene Pumpen
Lange vor der Erfindung der Photosynthese gab es schon Bakterien, die Energie aus der Oxidation von anorganischen Verbindungen gewinnen konnten. Schwefelbakterien beispielsweise oxidieren Schwefelwasserstoff H2S und andere reduzierte Schwefelverbindungen wie Thiosulfat S2O32- zu elementarem Schwefel oder Sulfat [1]. Die hierbei gewonnene Energie wird dann zum Antreiben einer Protonenpumpe verwendet. So wird ein Protonengradient erzeugt, der dann wiederum zur ATP-Gewinnung benutzt werden kann.
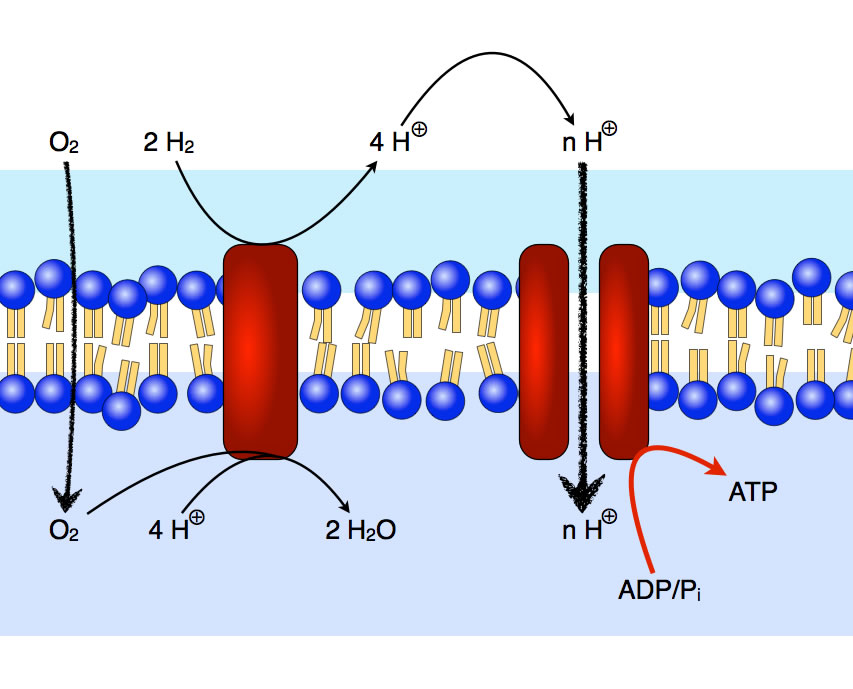
ATP-Erzeugung durch ein H2-oxidierendes Bakterium
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Das Wasserstoff oxidierende Bakterium Aquifex aeolicus oxidiert Wasserstoff zu Wasser und gewinnt dabei Energie, um auf indirektem Wege ATP zu erzeugen [2].
Der dafür erforderliche Protonengradient wird nicht durch den aktiven Transport von Protonen erzeugt, sondern durch die Freisetzung von Protonen auf der Außenseite der Membran und durch den gleichzeitigen Verbrauch von Protonen auf der Innenseite der Membran. Also durch zwei chemische Reaktionen, die - getrennt durch die Membran - unabhängig voneinander ablaufen. Der Sauerstoff, der zu "Entfernung" der Protonen auf der Innenseite benötigt wird, ist ein kleines unpolares Molekül und kann einfach durch die Lipid-Doppelschicht hindurch diffundieren.
Quellen:
- Alberts, Bruce et al. Molekularbiologie der Zelle, 6. Auflage, Weinheim 2017.
- Wikipedia, Artikel "Chemotrophie".
- Harvey Lodish et al. Molecular Cell Biology, New York 2004