Allgemeines
Die Abschlussgewebe der Pflanzen sind durchaus mit der Haut des Menschen vergleichbar: Sie grenzen den Pflanzenkörper nach außen ab und bieten sowohl einen mechanischen wie auch einen physiologischen bzw. chemischen Schutz. Wenn dennoch Stoffe aus der Umwelt aufgenommen oder an die Umwelt abgegeben werden müssen, dienen dazu spezielle Vorrichtungen der Abschlussgewebe, beispielsweise die Spaltöffnungen in der unteren Epidermis der Laubblätter.
Typisch für Abschlussgewebe ist, dass die Zellen sehr eng beieinander liegen, es sind so gut wie keine Interzellularen vorhanden. Weiterhin typisch für Abschlussgewebe ist das Fehlen von Chloroplasten (mit wenigen Ausnahmen wie in den Schließzellen der Spaltöffnungen). Zwar gibt es Plastiden in den Zellen der Abschlussgewebe, doch handelt es sich hierbei vorwiegend um Leukoplasten und Chromoplasten. Chromoplasten finden sich hauptsächlich in der Epidermis von Blütenblättern oder Früchten, wodurch diese in allen möglichen Farben erscheinen [1].
Beim Menschen gibt es auch innerhalb des Körpers Abschlussgewebe, die beispielsweise als Epidermis die Speiseröhre, die Atemwege, den Magen und andere innere Organe auskleiden bzw. nach außen gegen andere Organe abgrenzen. So ist das auch bei Pflanzen; auch hier können einzelne Gewebe durch eine Endodermis voneinander abgegrenzt sein. So ist das Leitgewebe in den Blättern von einer Endodermis umgeben, wodurch es vom Parenchym abgegrenzt ist.
Epidermis
Die Epidermis der Pflanzen ist meistens ein einschichtiges Abschlussgewebe, das zum Beispiel die Ober- und Unterseite der Laubblätter überzieht. Die Epidermis hat folgende Hauptfunktionen [3]:
Funktionen:
- Schutz vor Austrocknung,
- mechanischer Schutz,
- Schutz vor Tierfraß und Befall durch Parasiten,
- Ermöglichung eines geregelten und gezielten Gasaustausches,
- Sekretion bestimmter Stoffwechselprodukte,
- Absorption von Wasser,
- Bereitstellung von Sinneszellen für optische (Licht) und mechanische Reize.
Zelltypen:
Die Epidermis der Blätter enthält drei verschiedene Zelltypen.
- Normale Epidermiszellen
Diese Zellen stellen das Grundgerüst der Epidermis und bilden die eigentliche Abschluss- bzw. Schutzschicht. - Schließzellen
Jeweils zwei dieser Zellen bilden die Spaltöffnungen (Stomata). Mit diesen Spaltöffnungen können die Blätter Kohlendioxid aufnehmen und Wasserdampf abgeben. - Haarzellen
Pflanzenhaare können dem Schutz vor Fressfeinden dienen, manche enthalten sogar ätzende Verbindungen oder Giftstoffe oder sind sehr spitz. Oft dienen Pflanzenhaare aber auch dem Schutz vor Verdunstung, vor allem an den Spaltöffnungen mancher Pflanzen finden sich viele Haare.
Die typische Epidermiszelle besitzt unterschiedlich dicke Zellwände. Und zwar ist diejenige Seite der Zellwand, die nach außen zeigt, durch Sekundärwandschichten besonders stark ausgebildet, während die Zellwände, die Nachbarzellen voneinander trennen, eine normale Dicke besitzen. Allerdings sind die Zellwände, die benachbarte Epidermiszellen trennen, oft gewellt oder zackig geformt. Dadurch können sich benachbarte Epidermiszellen besser verzahnen, was die mechanische Festigkeit enorm erhöht.
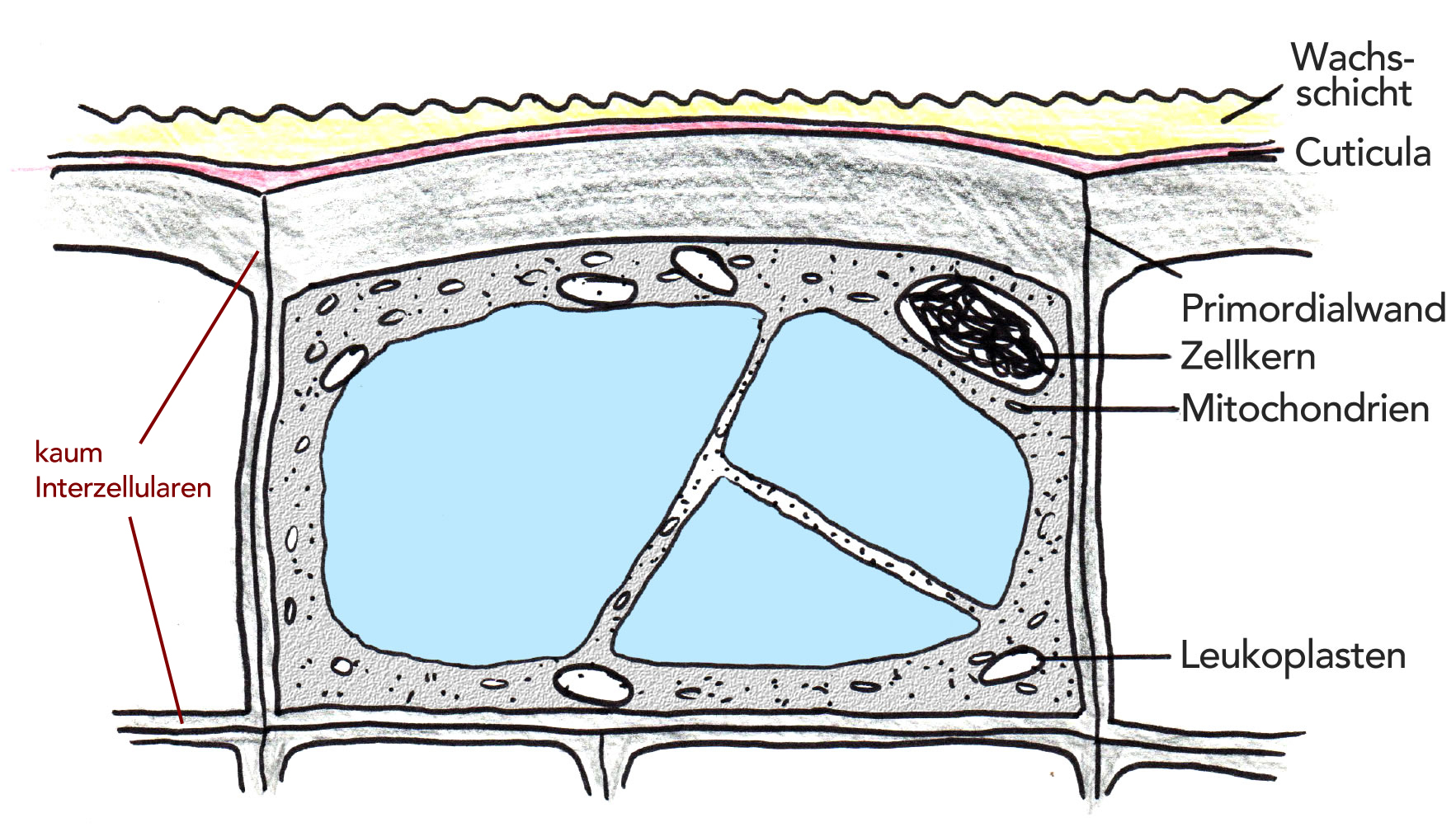
Eine typische Epidermiszelle
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Die Epidermiszellen der Angiospermen sind relativ chloroplastenfrei, es finden sich höchstens einige Leukoplasten, und in der Epidermis von Blütenblättern und Früchten auch Chromoplasten, die den Blüten und Früchten ihre Farbigkeit verleihen, um Insekten oder andere Tiere anzulocken.
Mono- und Dicotyledonen (Ein- und Zweikeimblättrige) besitzen unterschiedlich geformte Epidermiszellen mit charakteristischen Strukturen:
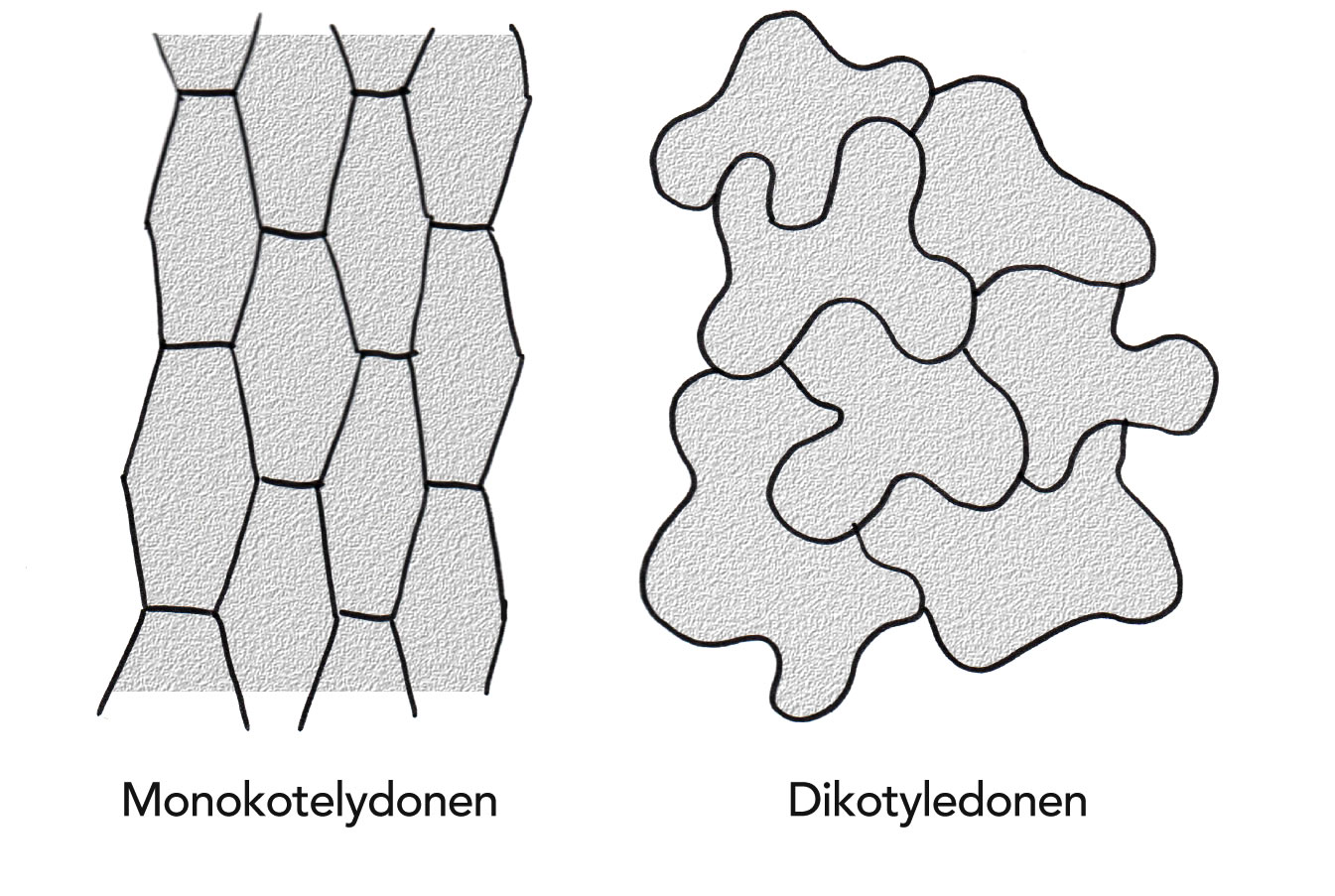
Aufsicht auf die Epidermen einer einkeimblättrigen (links) und einer zweikeimblättrigen (rechts) Pflanze
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Cuticula
Die Cuticula ist eine Schutzschicht, die von den Epidermiszellen gebildet wird, um das Blatt vor Wasser- und Gasverlust zu schützen.
Normale Zellwände sind recht gut durchlässig für Wasser und Gase, sie stellen in dieser Hinsicht also keinen effektiven Schutz dar. Aus diesem Grund haben die meisten Blätter Epidermiszellen mit besonders stark verdickten Außenzellwänden.
Diese Verdickung der äußeren Zellwände wird durch eine chemische Verbindung mit dem Namen Cutin bewirkt. Das Cutin wird vom Cytoplasma der Epidermiszellen hergestellt und durch die Zellwand nach außen abgegeben. Eine derart cutinisierte Zellwand ist nicht mehr so durchlässig für Wasser und Gase wie eine normale Zellwand. Setzt man die Verdunstungsrate einer freien Wasserfläche auf den Wert 100, hat die Verdunstungsrate durch eine Cuticula nur den Wert von 0,01 [1].
Die Cutinisierung der Zellwände erhöht außerdem die mechanische Festigkeit der Epidermis. Andererseits muss die Cuticula einer Pflanze in der Lage sein, das Wachstum der Pflanze mitzumachen. In diesem Fall wird das Cutin-Netzwerk durch besondere extrazelluläre Enzyme, die Cutinasen, gezielt aufgelockert [1]. Auch einige Pilze, die Pflanzen befallen, besitzen solche Cutinasen, damit sie besser in den Pflanzenkörper eindringen können [7].
Zwischen der verdickten Zellwand der Epidermiszellen und der Cuticula befindet sich eine sehr dünne, im Lichtmikroskop nicht sichtbare Schicht aus Protopektin[3].
Die Blattunterseiten und Wasserpflanzen besitzen in der Regel keine Cuticula und erst recht keine Wachsschicht, die einen weiteren Verdunstungsschutz für die Pflanze darstellt.
Pflanzen trockener Standorte hat oft eine sehr dicke Cuticula mit einer dicken Wachsschicht; bei Kakteen kann man das gut sehen. Manchmal wird die Cuticula auch durch Kalkabscheidungen oder Kieselsäureablagerungen verstärkt. Der Ackerschachtelhalm wurde früher als Zinnkraut bezeichnet, weil er früher zum Reinigen von Zinnbechern benutzt wurde. Durch seine Kieselsäureschicht eignete es sich dafür sehr gut [1, 9].
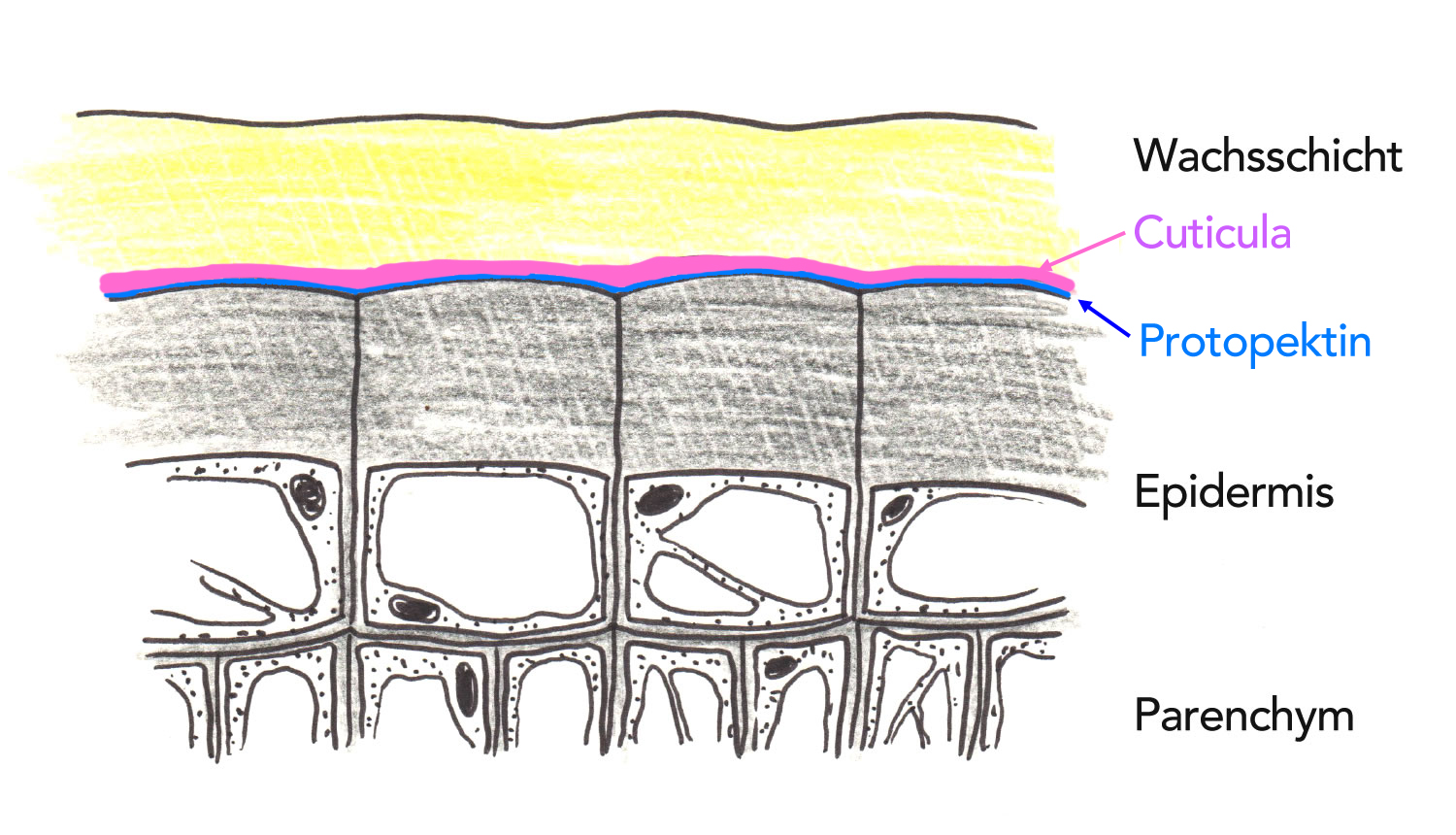
Die Epidermis eines Xerophyten
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Wachsschicht
In die cutinisierten Außenschichten der Zellwand ist oft auch Wachs eingelagert, welches die Zellwand noch undurchlässiger für Wasser und Gase macht. Manchmal ist diese Wachsschicht so dick, dass man sie schon mit bloßem Auge sehen kann. Auf Obst (z.B. bei Pflaumen) bildet sie einen hellgrauen, abwischbaren Reif.
Wachse sind chemisch gesehen Ester aus einem langkettigen Alkohol und einer langkettigen Fettsäure. Eine solche Wachsschicht kann, wenn sie besonders dick ist, schon mit dem bloßen Auge als weißlicher Belag auf dem Blatt oder dem Stängel wahrgenommen werden (Wachsreif). Durch einen Wachsüberzug kann ein Blatt völlig unbenetzbar werden, Wassertropfen rollen ungehindert einfach ab. In der Technik wird ein solcher Effekt als Lotuseffekt bezeichnet. Die Wachsmoleküle werden in der Epithelzelle hergestellt und gelangen mit dem wenigen Wasser, das die Cuticula passiert, in die Wachsschicht [1].
Die Cuticula und die Wachsschicht waren offensichtlich eine "Erfindung" der Pflanzen, als sie das Land eroberten. Die wachsüberzogene Cuticula verhindert nicht nur den Wasserverlust, sondern stabilisiert auch die Blätter und Stängel, schützt sie also etwas vor der Schwerkraft, die an Land ja viel höher ist als im Wasser [8].
Spaltöffnungen der unteren Blatt-Epidermis
Epidermis mit Spaltöffnungen auf der Blattunterseite von Helleborus niger, Vergrößerung 120x
Autor: Strasburger 1906, neu gezeichnet von U. Helmich 2021
Diese Zeichnung stammt aus dem berühmten Strasburger, der "Bibel der Botaniker", der 8. Auflage aus dem Jahre 1906.
Eduard Strasburger (1844 - 1912) hat nicht nur das "Lehrbuch für Botanik für Hochschulen" geschrieben (1. Auflage 1894), sondern hat auch 1875 die Teilung des pflanzlichen Zellkerns entdeckt, war also auch ein bekannter Wissenschaftler. Das Lehrbuch der Botanik für Hochschulen gibt es immer noch, es heißt jetzt allerdings "Lehrbuch der Pflanzenwissenschaften" und ist 2021 in der 38. Auflage erschienen [1].
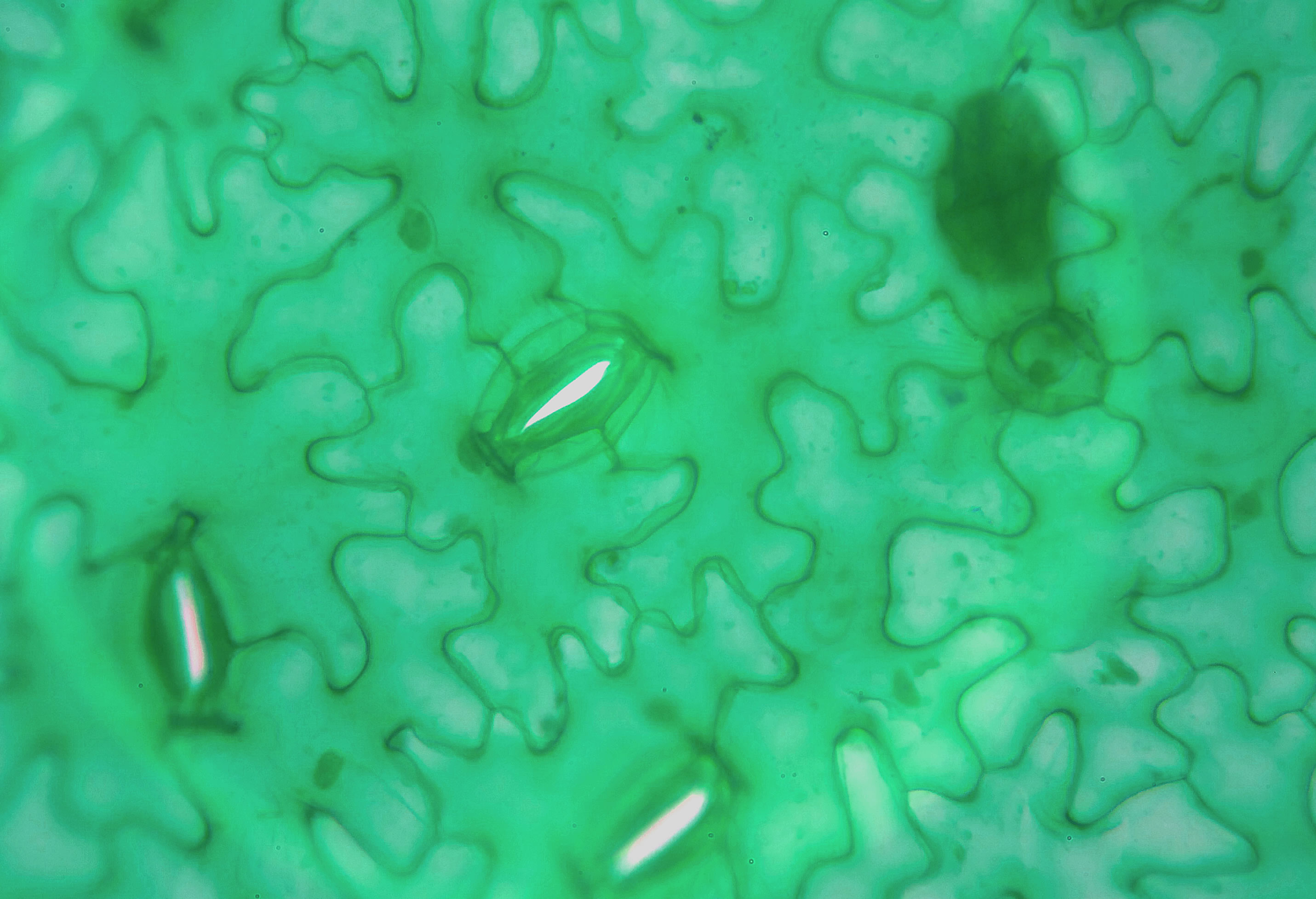
Spaltöffnungen in einem Laubblatt
Photo: Ulrich Helmich 2022, Lizenz: Public domain
Dieses Photo habe ich von einem Fertig-Mikropräparat gemacht; die Pflanzenart stand leider nicht auf dem Etikett des Objektträgers.
Technisches
Labormikroskop der Fa. Christoph Beck (Kassel, gibt es leider seit 1982 nicht mehr), Objektiv 40x, Beleuchtung mit LED-Videolicht, Kameraadapter von Bresser, Canon 5D Mark II, Remote-Aufnahme mit Canon EOS Utility-Software, Bild aus fünf Photos zusammengestackt (Photostacking mit der MacOS-App Focus Stacker).
Die Technik habe ich hier so ausführlich beschrieben, weil es das erste Mikrophoto ist, das ich nach diesem Verfahren (Kamera-Adapter, Focus-Stacking) erstellt habe.
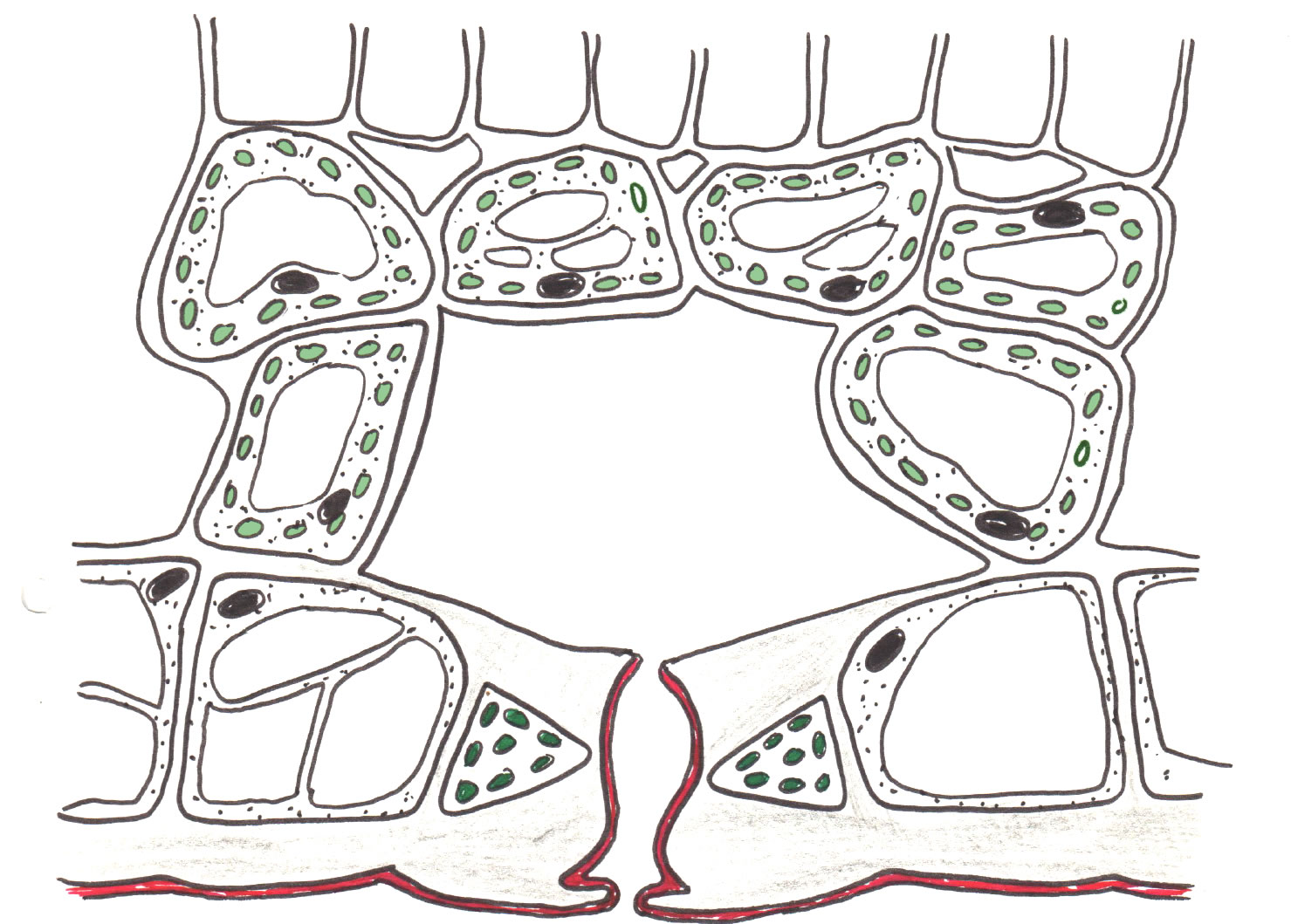
Spaltöffnung in der unteren Epidermis eines Laubblattes
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Hier sehen wir einen Querschnitt durch den unteren Teil eines Laubblattes mit einer Spaltöffnung. Die Cuticula ist wieder rot hervorgehoben, eine Wachsschicht findet sich auf der unteren Epidermis in der Regel nicht. Man achte darauf, dass die Zellen der unteren (und oberen) Epidermis keine Chloroplasten enthalten, wohl aber die Schließzellen der Spaltöffnungen.
Die Öffnung ist abhängig von Umweltfaktoren
Photonastie
Sowohl Rotlicht wie auch Blaulicht führen zu einer Öffnung der Spaltöffnungen. Blaulicht wirkt allerdings stärker, weil es direkt wirkt (siehe Öffnungsmechanismus). Rotlicht wirkt indirekt auf die Spaltöffnungsweite über die Photosynthese des Blattes und der damit verbundenen ATP-Erzeugung sowie der Reduktion der CO2-Konzentration.
Thermonastie
Eine Erhöhung der Temperatur beschleunigt nach der RGT-Regel biochemische Prozesse. Das Öffnen der Stomata sollte also eigentlich schneller gehen, wenn die Temperaturen steigen. Allerdings droht bei höheren Temperaturen auch immer die Gefahr eines Wasserverlustes durch zu starke Transpiration. Um diesen Wasserverlust zu verhindern, schließen sich die Stomata bei erhöhten Temperaturen.
Chemonastie
Die CO2-Konzentration in den Interzellularen hat einen ganz entscheidenden Einfluss auf die Öffnungsweite der Stomata. Bei niedriger CO2-Konzentration öffnen sich die Stomata, damit die Pflanze leichter an Kohlendioxid gelangen kann, das sie ja für die Photosynthese benötigt. Bei hoher CO2-Konzentration sind die Stomata eher geschlossen. Die chemischen CO2-Sensoren befinden sich wahrscheinlich in den Schließzellen selbst, wie die CO2-gesteuerte Regulation der Spaltöffnungsweite aber genau funktioniert, ist noch größtenteils ungeklärt.
Hydronastie
Bei Wassermangel wird das Phytohormon Abscisinsäure produziert, das einen schnellen Spaltenschluss der Stomata bewirkt. Die genaue Wirkweise von Abscisinsäure ist auf der Lexikonseite "Spaltöffnungen, Mechanismus" erläutert.
Öffnungsmechanismus
Durch einen Turgormechanismus werden die Spaltöffnungen in der unteren Epidermis eines Blattes reguliert. Bei hoher Luftfeuchtigkeit sind die Schließzellen und ihre Nebenzellen reich an Wasser, der Turgor ist also hoch und die dünne Zellwand an der Rückseite der Schließzelle kann sich ausdehnen. Die dicken Zellwände der Schließzelle dagegen können sich nicht selbst ausdehnen, sondern müssen die Bewegung der dünnen, dehnbaren Zellwand mitmachen. Dadurch wird der Zentralspalt größer, und die Pflanze kann leichter Wasser abgeben.
Auf dieser Seite finden Sie mehr Informationen zu den Spaltöffnungen. Vor allem wird hier auch der Mechanismus erläutert, der dazu führt, dass sich die Stomata öffnen, wenn beispielsweise Licht auf das Blatt fällt.
Entstehung
Spaltöffnungen entstehen durch inäquale Teilung normaler Epidermiszellen, die ihre Teilungsfähigkeit wiedererlangt haben (Meristemoide).
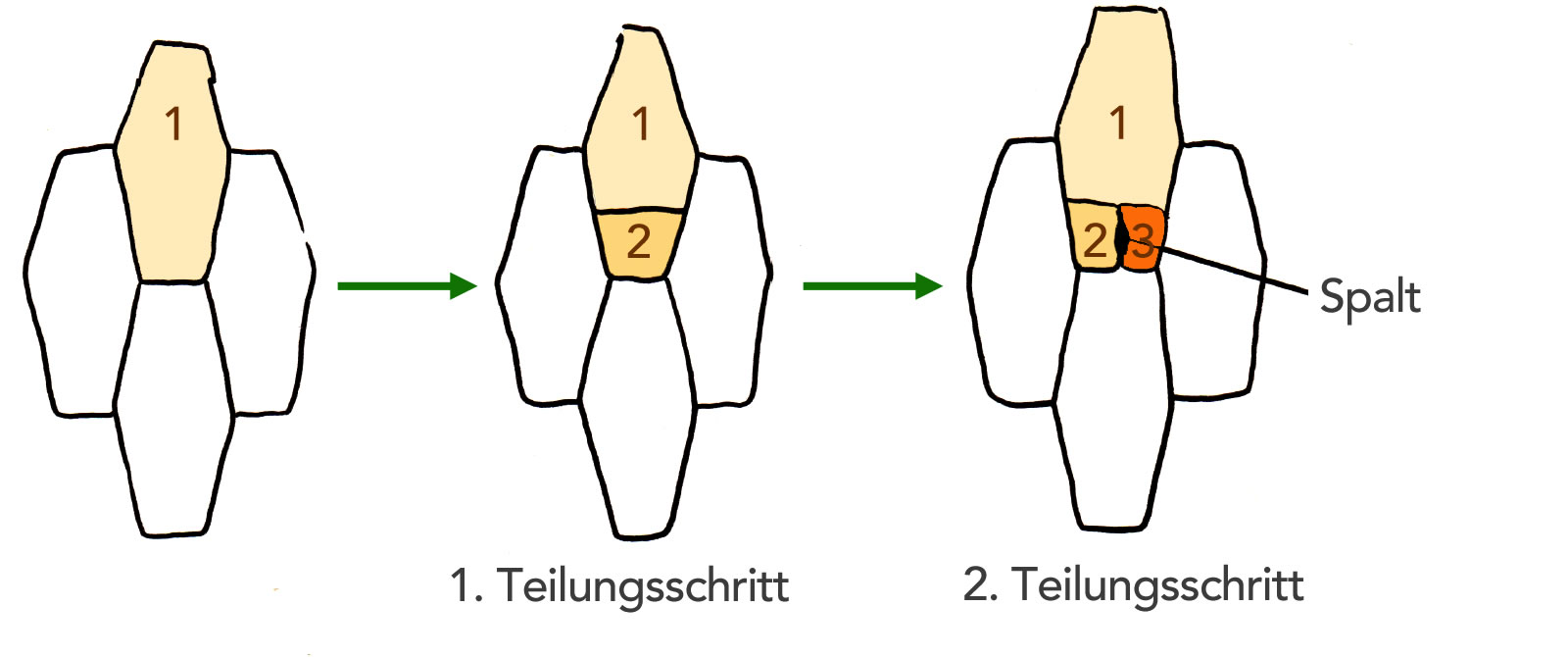
Bildung einer Spaltöffnung
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Typen
Außerdem bisher besprochenen Amaryllen-Typ gibt es noch zwei andere wichtige und weit verbreitete Typen von Spaltöffnungen.
In diesem Abschnitt auf der Lexikonseite "Spaltöffnungen" finden Sie weitere Informationen zu den verschiedenen Typen der Spaltöffnungen.
Haare
Zahlreiche Epidermen bringen besondere Haare oder Trichome hervor. Diese entstehen stets aus einer einzigen Epidermiszelle, einer sogenannten Initialzelle. Pflanzenhaare sind somit stets in der Epidermis verankert. Sie können vielfältige Funktionen haben:
- Wurzelhaare dienen der Aufnahme von Wasser mit gelösten Nährsalzen.
- Frucht- und Samenhaare erleichtern die Ausbreitung von Früchten und Samen mit dem Wind. Ein besonders extremes Beispiel für Samenhaare sind die Haare der Baumwollsamen, die bis zu 5 cm lang werden und die Grundlage einer ganzen Industrie darstellen.
- Nebelhaare an Stängeln oder Blättern dienen zur Aufnahme von Wasser in Gegenden, wo es oft nebelig ist.
- Einige tote und luftgefüllte Haare dienen als Strahlenschutz an sehr sonnigen Standorten.
- Klimmhaare dienen zum Festhalten der Pflanze am Untergrund, an Steinen, Zweigen, Baumstämmen oder anderen Objekten der Umgebung. Man denke nur an das Klettenlabkraut oder an Wein.
- Borstenhaare mit verkieselten Zellwänden sind sehr hart und spitz und dienen zur Feindabwehr,
- Brennhaare sind mit ätzenden Flüssigkeiten gefüllt und bewirken eine noch effektivere Feindabwehr.
- Fühlhaare, zum Beispiel bei der Venusfliegenfalle, dienen zum Erkennen von Beutetieren (Insekten).
- Drüsenhaare produzieren chemische Verbindungen, die leicht abgegeben werden können (Schutz vor Fraßfeinden)

Bildung eines Pflanzenhaars
Autor: Ulrich Helmich 1978/2021, Lizenz: Public domain
Diese Haare können einzellig oder mehrzellig, lebend oder abgestorben sein.
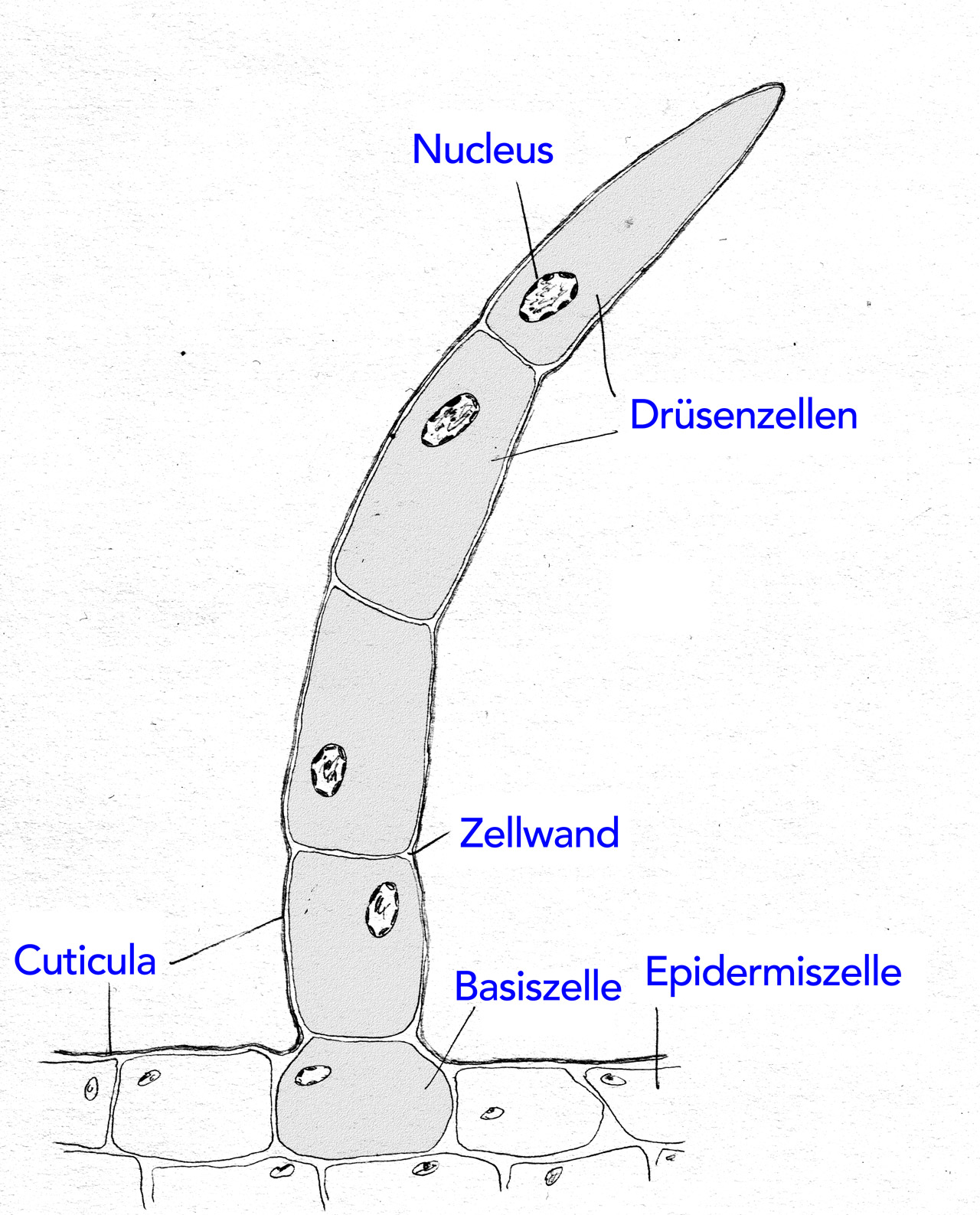
Drüsenhaar von Coleus (Buntnessel, Lippenblütler)
Autor: Ulrich Helmich 1979/2021, Lizenz: Public domain
Hier sehen wir ein mehrzelliges Drüsenhaar einer Buntnessel.
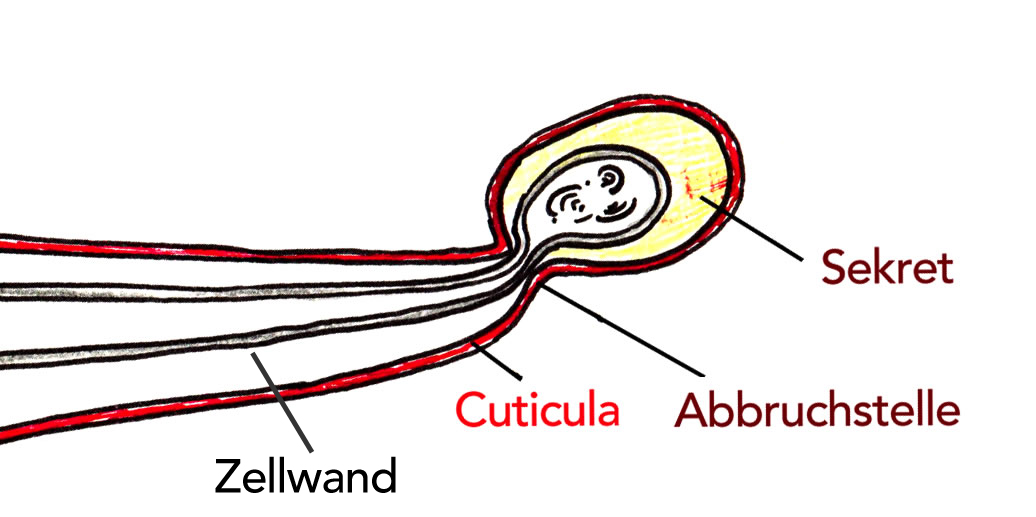
Brennhaar einer Brennnessel mit einer Sollbruchstelle
Autor: Ulrich Helmich 2021, Lizenz: Public domain
Eine besondere Funktion erfüllen die Brennhaare einiger Pflanzen. Sie dienen der Abwehr von Fressfeinden. Zwischen Cuticula und Zellwand ist ein Zwischenraum, in dem sich Sekrete der lebenden Zelle ansammeln. Bei der geringsten Berührung bricht das Köpfchen ab, und die Sekrete (Ameisensäure, Acetylcholin und Histidin) treten aus.
Endodermis
Eine "innere" Epidermis trennt Gewebekomplexe voneinander und fungiert als physiologische Barriere. Am häufigsten trifft man eine Endodermis in den Wurzeln der Kormophyten (Farnpflanzen und Samenpflanzen) an.
Die Endodermis trennt dort den Zentralzylinder von der Wurzelrinde und ist eine physiologische Barriere für den Übertritt von Wasser, das von den Wurzelhaaren aus dem Boden aufgenommen wurde, aus der Rinde in den Zentralzylinder.
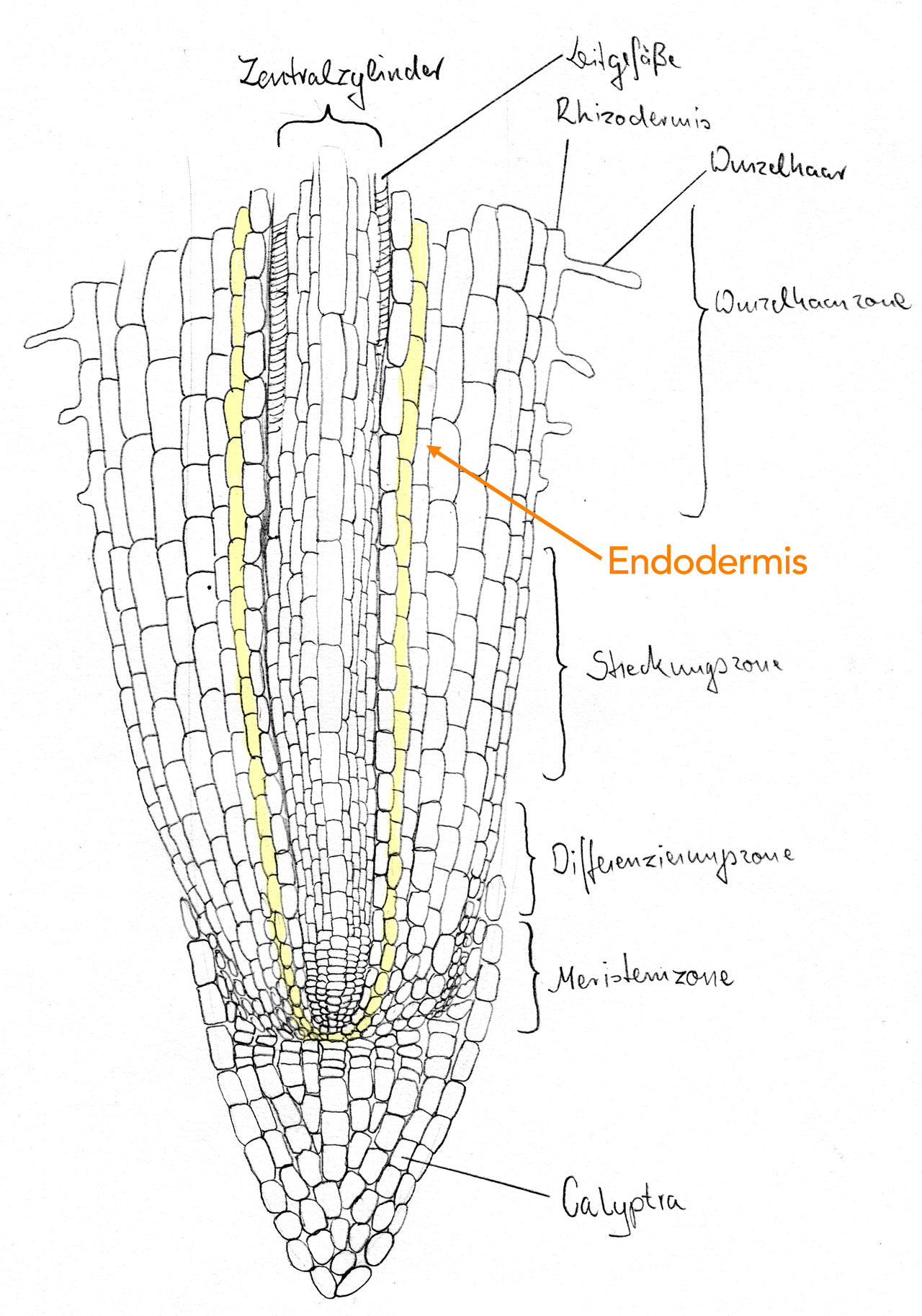
Wurzel eines Grases (Poa nemoralis)
Autor: Ulrich Helmich 1979/2021, Lizenz: Public domain
Die Endodermiszellen sind für diesen Zweck mit einem die Radialwände umlaufenden Suberinband, dem Casparischen Streifen, ausgerüstet. Dieser ist wasserundurchlässig und verschließt die Kapillarräume zwischen den Zellwänden benachbarter Zellen. Dadurch kann kein Wasser durch diese Kapillarräume transportiert werden, sondern der Transport muss durch den Protoplasten der Endodermiszellen selber bewerkstelligt werden. Der Vorteil liegt darin, dass dieser Transport im Gegensatz zum kapillaren Transport besser reguliert werden kann.
Quellen und allgemeine Literatur zur Botanik:
- Kadereit , Körner, Nick, Sonnewald: Strasburger - Lehrbuch der Pflanzenwissenschaften, 38. Auflage, Springer Berlin Heidelberg 2021.
- Wilhelm Nultsch: Allgemeine Botanik, 7. Auflage, Stuttgart 1982
- Wilhelm Nultsch: Allgemeine Botanik, 11. Auflage, Stuttgart 2001
- Urry, Cain, Wassermann, Minorsky, Reece. Campbell Biologie, Hallbergmoos 2019, 11.Auflage.
- Savada, Hillis, Heller, Hacker: Purves Biologie, Springer Verlag Deutschland 2019, 10. Auflage. Herausgegeben von Jürgen Markl.
- engl. Wikipedia, Artikel "Cutin".
- Wikipedia, Artikel "Cutinase".
- "Schutzschild der Pflanzen" auf Pflanzenforschung.de
- Wikipedia, Artikel "Ackerschachtelhalm".